

Integrin adhesion drives the emergent polarization of active cytoskeletal stresses to pattern cell delamination
- PMID: 21571643
- PMCID: PMC3107263
- DOI: 10.1073/pnas.1018652108
Integrin adhesion drives the emergent polarization of active cytoskeletal stresses to pattern cell delamination
Abstract
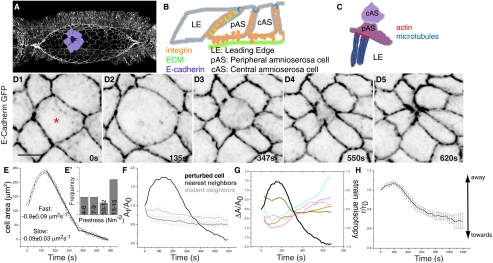
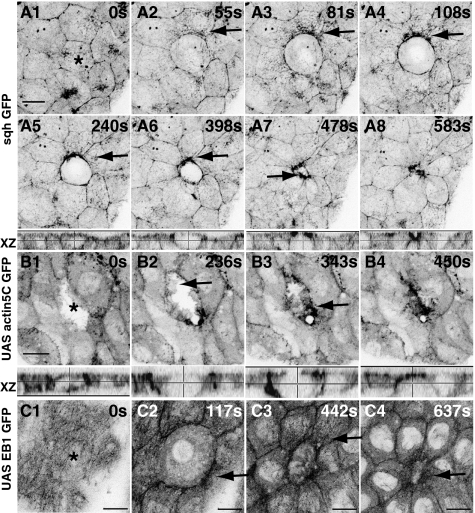
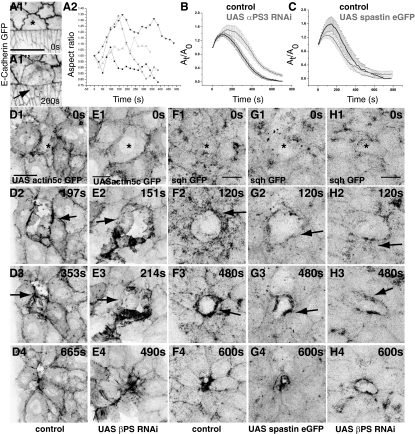
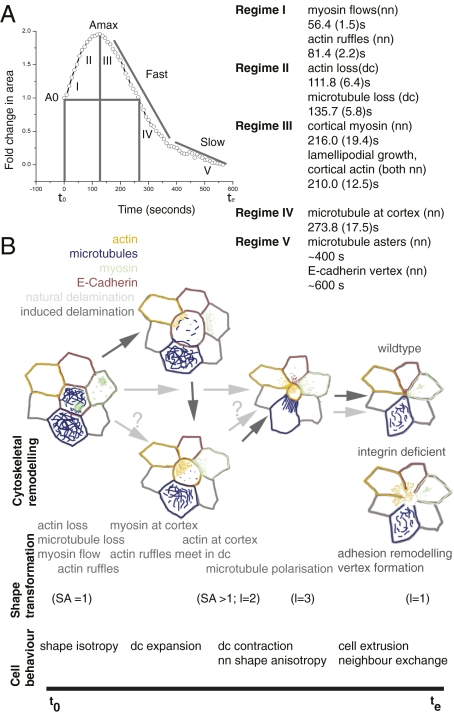
Tissue patterning relies on cellular reorganization through the interplay between signaling pathways and mechanical stresses. Their integration and spatiotemporal coordination remain poorly understood. Here we investigate the mechanisms driving the dynamics of cell delamination, diversely deployed to extrude dead cells or specify distinct cell fates. We show that a local mechanical stimulus (subcellular laser perturbation) releases cellular prestress and triggers cell delamination in the amnioserosa during Drosophila dorsal closure, which, like spontaneous delamination, results in the rearrangement of nearest neighbors around the delaminating cell into a rosette. We demonstrate that a sequence of "emergent cytoskeletal polarities" in the nearest neighbors (directed myosin flows, lamellipodial growth, polarized actomyosin collars, microtubule asters), triggered by the mechanical stimulus and dependent on integrin adhesion, generate active stresses that drive delamination. We interpret these patterns in the language of active gels as asters formed by active force dipoles involving surface and body stresses generated by each cell and liken delamination to mechanical yielding that ensues when these stresses exceed a threshold. We suggest that differential contributions of adhesion, cytoskeletal, and external stresses must underlie differences in spatial pattern.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Spatial, temporal and molecular hierarchies in the link between death, delamination and dorsal closure.Development. 2011 Jul;138(14):3043-54. doi: 10.1242/dev.060731. Development. 2011. PMID: 21693520
-
A Cdc42-mediated supracellular network drives polarized forces and Drosophila egg chamber extension.Nat Commun. 2020 Apr 21;11(1):1921. doi: 10.1038/s41467-020-15593-2. Nat Commun. 2020. PMID: 32317641 Free PMC article.
-
Mitochondrial ROS regulates cytoskeletal and mitochondrial remodeling to tune cell and tissue dynamics in a model for wound healing.Dev Cell. 2014 Feb 10;28(3):239-52. doi: 10.1016/j.devcel.2013.12.019. Epub 2014 Jan 30. Dev Cell. 2014. PMID: 24486154
-
Signaling pathways directing the movement and fusion of epithelial sheets: lessons from dorsal closure in Drosophila.Differentiation. 2002 Jun;70(4-5):181-203. doi: 10.1046/j.1432-0436.2002.700408.x. Differentiation. 2002. PMID: 12147138 Review.
-
Cell adhesion in the assembly of the Drosophila eye.J Neurogenet. 2014 Sep-Dec;28(3-4):282-90. doi: 10.3109/01677063.2014.907799. Epub 2014 May 28. J Neurogenet. 2014. PMID: 24678921 Review.
Cited by
-
Two types of critical cell density for mechanical elimination of abnormal cell clusters from epithelial tissue.PLoS Comput Biol. 2022 Jun 13;18(6):e1010178. doi: 10.1371/journal.pcbi.1010178. eCollection 2022 Jun. PLoS Comput Biol. 2022. PMID: 35696420 Free PMC article.
-
Cell crawling mediates collective cell migration to close undamaged epithelial gaps.Proc Natl Acad Sci U S A. 2012 Jul 3;109(27):10891-6. doi: 10.1073/pnas.1117814109. Epub 2012 Jun 18. Proc Natl Acad Sci U S A. 2012. PMID: 22711834 Free PMC article.
-
Local, cell-nonautonomous feedback regulation of myosin dynamics patterns transitions in cell behavior: a role for tension and geometry?Mol Biol Cell. 2013 Aug;24(15):2350-61. doi: 10.1091/mbc.E12-12-0868. Epub 2013 Jun 5. Mol Biol Cell. 2013. PMID: 23741052 Free PMC article.
-
Theoretical Analysis of Stress Distribution and Cell Polarization Surrounding a Model Wound.Biophys J. 2018 Jul 17;115(2):398-410. doi: 10.1016/j.bpj.2018.06.011. Biophys J. 2018. PMID: 30021114 Free PMC article.
-
Unconventional Ways to Live and Die: Cell Death and Survival in Development, Homeostasis, and Disease.Annu Rev Cell Dev Biol. 2018 Oct 6;34:311-332. doi: 10.1146/annurev-cellbio-100616-060748. Epub 2018 Aug 8. Annu Rev Cell Dev Biol. 2018. PMID: 30089222 Free PMC article. Review.
References
-
- Lecuit T. Adhesion remodeling underlying tissue morphogenesis. Trends Cell Biol. 2005;15:34–42. - PubMed
-
- Juliano RL. Signal transduction by cell adhesion receptors and the cytoskeleton: Functions of integrins, cadherins, selectins, and immunoglobulin-superfamily members. Annu Rev Pharmacol Toxicol. 2002;42:283–323. - PubMed
-
- Bershadsky AD, Balaban NQ, Geiger B. Adhesion-dependent cell mechanosensitivity. Annu Rev Cell Dev Biol. 2003;19:677–695. - PubMed
-
- Vogel V, Sheetz M. Local force and geometry sensing regulate cell functions. Nat Rev Mol Cell Biol. 2006;7:265–275. - PubMed
-
- Hutson MS, et al. Forces for morphogenesis investigated with laser microsurgery and quantitative modeling. Science. 2003;300:145–149. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
