

Targeted mutagenesis of duplicated genes in soybean with zinc-finger nucleases
- PMID: 21464476
- PMCID: PMC3177250
- DOI: 10.1104/pp.111.172981
Targeted mutagenesis of duplicated genes in soybean with zinc-finger nucleases
Abstract
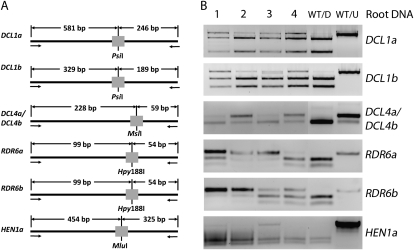
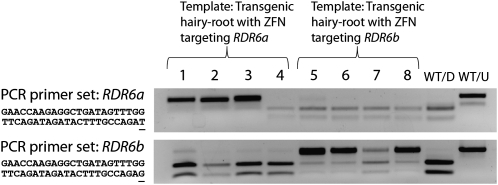
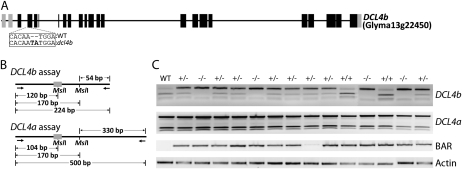
We performed targeted mutagenesis of a transgene and nine endogenous soybean (Glycine max) genes using zinc-finger nucleases (ZFNs). A suite of ZFNs were engineered by the recently described context-dependent assembly platform--a rapid, open-source method for generating zinc-finger arrays. Specific ZFNs targeting dicer-like (DCL) genes and other genes involved in RNA silencing were cloned into a vector under an estrogen-inducible promoter. A hairy-root transformation system was employed to investigate the efficiency of ZFN mutagenesis at each target locus. Transgenic roots exhibited somatic mutations localized at the ZFN target sites for seven out of nine targeted genes. We next introduced a ZFN into soybean via whole-plant transformation and generated independent mutations in the paralogous genes DCL4a and DCL4b. The dcl4b mutation showed efficient heritable transmission of the ZFN-induced mutation in the subsequent generation. These findings indicate that ZFN-based mutagenesis provides an efficient method for making mutations in duplicate genes that are otherwise difficult to study due to redundancy. We also developed a publicly accessible Web-based tool to identify sites suitable for engineering context-dependent assembly ZFNs in the soybean genome.
Figures


Similar articles
-
Rapid mutation of endogenous zebrafish genes using zinc finger nucleases made by Oligomerized Pool ENgineering (OPEN).PLoS One. 2009;4(2):e4348. doi: 10.1371/journal.pone.0004348. Epub 2009 Feb 9. PLoS One. 2009. PMID: 19198653 Free PMC article.
-
High frequency targeted mutagenesis in Arabidopsis thaliana using zinc finger nucleases.Proc Natl Acad Sci U S A. 2010 Jun 29;107(26):12028-33. doi: 10.1073/pnas.0914991107. Epub 2010 May 27. Proc Natl Acad Sci U S A. 2010. PMID: 20508152 Free PMC article.
-
Rapid "open-source" engineering of customized zinc-finger nucleases for highly efficient gene modification.Mol Cell. 2008 Jul 25;31(2):294-301. doi: 10.1016/j.molcel.2008.06.016. Mol Cell. 2008. PMID: 18657511 Free PMC article.
-
[The mechanism and application of zinc finger nucleases].Yi Chuan. 2011 Feb;33(2):123-30. doi: 10.3724/sp.j.1005.2011.00123. Yi Chuan. 2011. PMID: 21377968 Review. Chinese.
-
Advances in genetic modification of farm animals using zinc-finger nucleases (ZFN).Chromosome Res. 2015 Feb;23(1):7-15. doi: 10.1007/s10577-014-9451-7. Chromosome Res. 2015. PMID: 25596823 Review.
Cited by
-
Direct stacking of sequence-specific nuclease-induced mutations to produce high oleic and low linolenic soybean oil.BMC Plant Biol. 2016 Oct 13;16(1):225. doi: 10.1186/s12870-016-0906-1. BMC Plant Biol. 2016. PMID: 27733139 Free PMC article.
-
Genetically modified (GM) crops: milestones and new advances in crop improvement.Theor Appl Genet. 2016 Sep;129(9):1639-55. doi: 10.1007/s00122-016-2747-6. Epub 2016 Jul 5. Theor Appl Genet. 2016. PMID: 27381849 Review.
-
Genome Editing and Improvement of Abiotic Stress Tolerance in Crop Plants.Life (Basel). 2023 Jun 27;13(7):1456. doi: 10.3390/life13071456. Life (Basel). 2023. PMID: 37511831 Free PMC article. Review.
-
Advanced genetic tools for plant biotechnology.Nat Rev Genet. 2013 Nov;14(11):781-93. doi: 10.1038/nrg3583. Epub 2013 Oct 9. Nat Rev Genet. 2013. PMID: 24105275 Review.
-
Activities and specificities of homodimeric TALENs in Saccharomyces cerevisiae.Curr Genet. 2014 May;60(2):61-74. doi: 10.1007/s00294-013-0412-z. Epub 2013 Oct 1. Curr Genet. 2014. PMID: 24081604
References
-
- Curtin SJ, Watson JM, Smith NA, Eamens AL, Blanchard CL, Waterhouse PM. (2008) The roles of plant dsRNA-binding proteins in RNAi-like pathways. FEBS Lett 582: 2753–2760 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
