

Retromer terminates the generation of cAMP by internalized PTH receptors
- PMID: 21445058
- PMCID: PMC3079799
- DOI: 10.1038/nchembio.545
Retromer terminates the generation of cAMP by internalized PTH receptors
Abstract
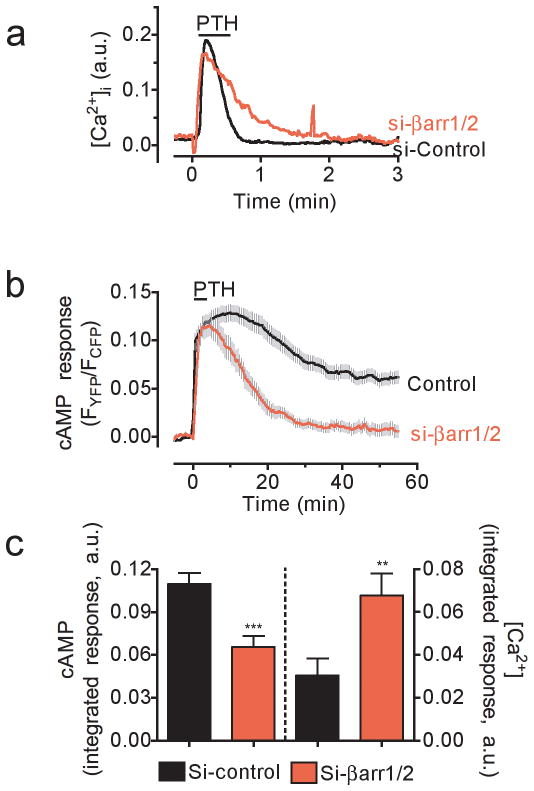
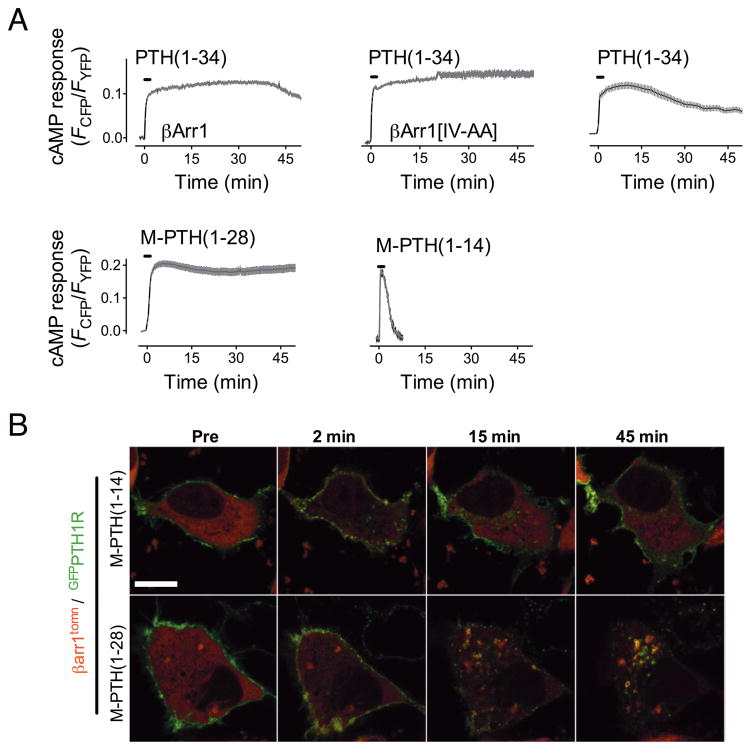
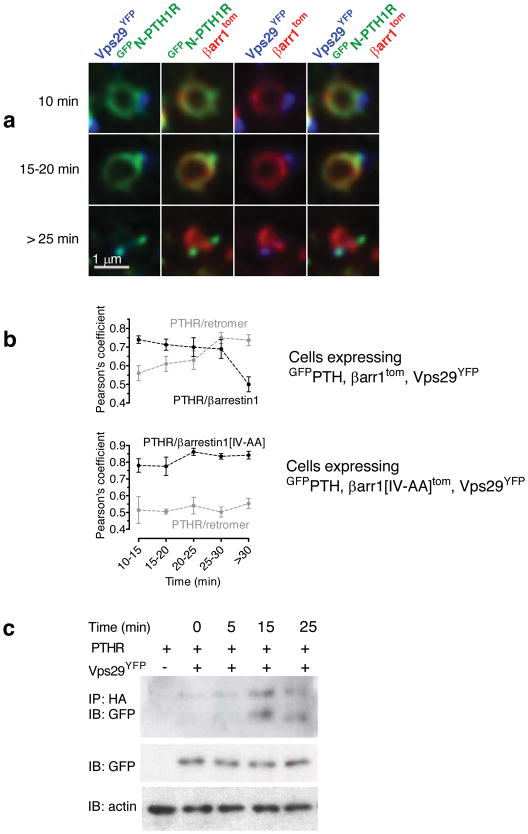
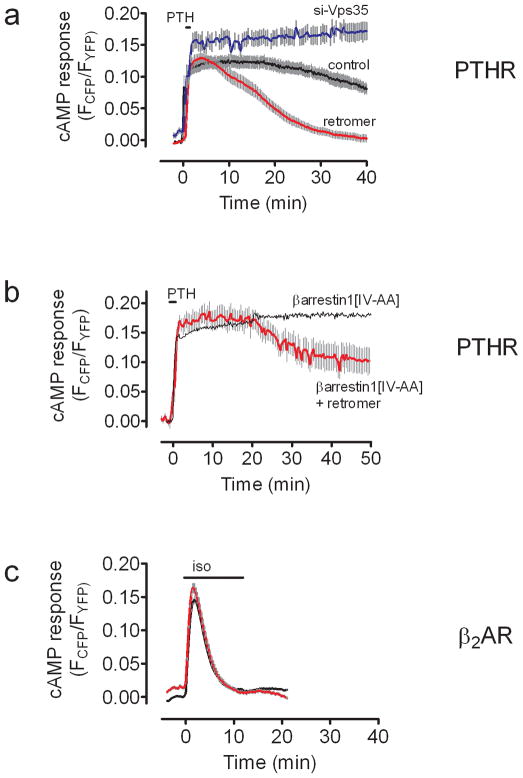
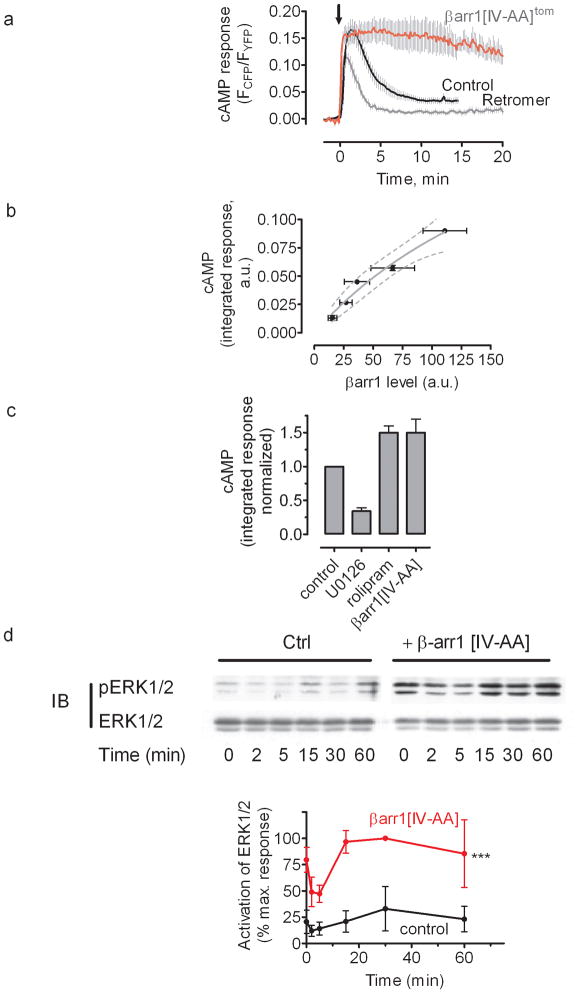
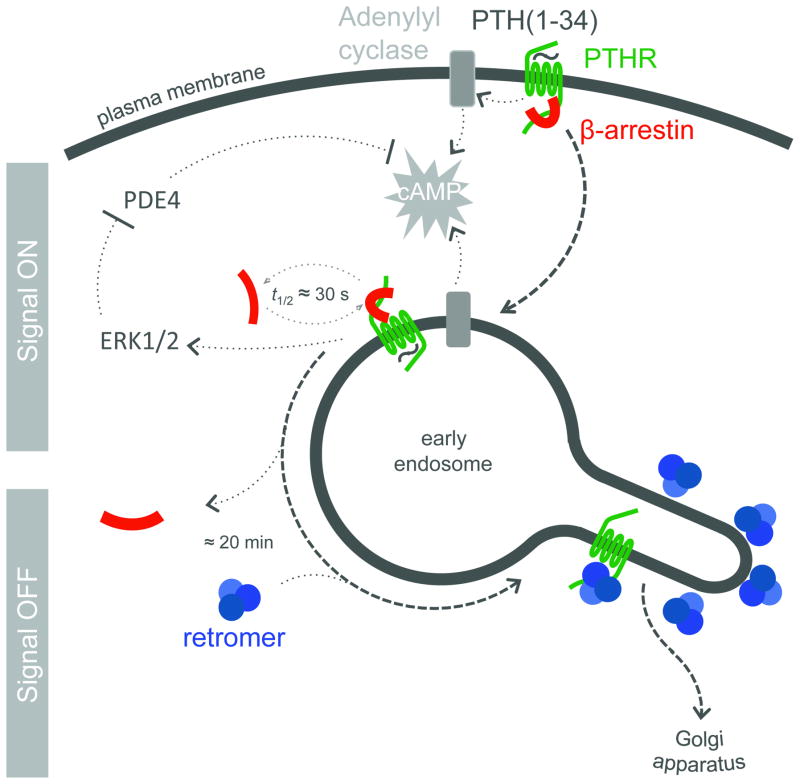
The generation of cAMP by G protein-coupled receptors (GPCRs) and its termination are currently thought to occur exclusively at the plasma membrane of cells. Under existing models of receptor regulation, this signal is primarily restricted by desensitization of the receptors through their binding to β-arrestins. However, this paradigm is not consistent with recent observations that the parathyroid hormone receptor type 1 (PTHR) continues to stimulate cAMP production even after receptor internalization, as β-arrestins are known to rapidly bind and internalize activated PTHR. Here we show that binding to β-arrestin1 prolongs rather than terminates the generation of cAMP by PTHR, and that cAMP generation correlates with the persistence of arrestin-receptor complexes on endosomes. PTHR signaling is instead turned off by the retromer complex, which regulates the movement of internalized receptor from endosomes to the Golgi apparatus. Thus, binding by the retromer complex regulates the sustained generation of cAMP triggered by an internalized GPCR.
Conflict of interest statement
The authors have no competing financial interests to disclose.
Figures
Comment in
-
Signaling: retromer arrests receptor on the run.Nat Chem Biol. 2011 May;7(5):251-2. doi: 10.1038/nchembio.564. Nat Chem Biol. 2011. PMID: 21502946 Free PMC article.
Similar articles
-
Noncanonical GPCR signaling arising from a PTH receptor-arrestin-Gβγ complex.Proc Natl Acad Sci U S A. 2013 Jan 22;110(4):1530-5. doi: 10.1073/pnas.1205756110. Epub 2013 Jan 7. Proc Natl Acad Sci U S A. 2013. PMID: 23297229 Free PMC article.
-
Endosomal parathyroid hormone receptor signaling.Am J Physiol Cell Physiol. 2022 Sep 1;323(3):C783-C790. doi: 10.1152/ajpcell.00452.2021. Epub 2022 Aug 1. Am J Physiol Cell Physiol. 2022. PMID: 35912987 Free PMC article. Review.
-
Gq/11-dependent regulation of endosomal cAMP generation by parathyroid hormone class B GPCR.Proc Natl Acad Sci U S A. 2020 Mar 31;117(13):7455-7460. doi: 10.1073/pnas.1918158117. Epub 2020 Mar 17. Proc Natl Acad Sci U S A. 2020. PMID: 32184323 Free PMC article.
-
Non-canonical signaling of the PTH receptor.Trends Pharmacol Sci. 2012 Aug;33(8):423-31. doi: 10.1016/j.tips.2012.05.004. Epub 2012 Jun 16. Trends Pharmacol Sci. 2012. PMID: 22709554 Free PMC article. Review.
-
Noncanonical control of vasopressin receptor type 2 signaling by retromer and arrestin.J Biol Chem. 2013 Sep 27;288(39):27849-60. doi: 10.1074/jbc.M112.445098. Epub 2013 Aug 9. J Biol Chem. 2013. PMID: 23935101 Free PMC article.
Cited by
-
Vps35 loss promotes hyperresorptive osteoclastogenesis and osteoporosis via sustained RANKL signaling.J Cell Biol. 2013 Mar 18;200(6):821-37. doi: 10.1083/jcb.201207154. J Cell Biol. 2013. PMID: 23509071 Free PMC article.
-
Identification of key phosphorylation sites in PTH1R that determine arrestin3 binding and fine-tune receptor signaling.Biochem J. 2016 Nov 15;473(22):4173-4192. doi: 10.1042/BCJ20160740. Epub 2016 Sep 13. Biochem J. 2016. PMID: 27623777 Free PMC article.
-
Therapeutic Targeting of Endosomal G-Protein-Coupled Receptors.Trends Pharmacol Sci. 2018 Oct;39(10):879-891. doi: 10.1016/j.tips.2018.08.003. Epub 2018 Sep 1. Trends Pharmacol Sci. 2018. PMID: 30180973 Free PMC article. Review.
-
Preassembled GPCR signaling complexes mediate distinct cellular responses to ultralow ligand concentrations.Sci Signal. 2018 Oct 9;11(551):eaan1188. doi: 10.1126/scisignal.aan1188. Sci Signal. 2018. PMID: 30301787 Free PMC article.
-
Regulation of Somatostatin Receptor 2 Trafficking by C-Tail Motifs and the Retromer.Endocrinology. 2019 May 1;160(5):1031-1043. doi: 10.1210/en.2018-00865. Endocrinology. 2019. PMID: 30822353 Free PMC article.
References
-
- Perry SJ, et al. Targeting of cyclic AMP degradation to beta 2-adrenergic receptors by beta-arrestins. Science. 2002;298:834–6. - PubMed
-
- Premont RT, Gainetdinov RR. Physiological roles of G protein-coupled receptor kinases and arrestins. Annu Rev Physiol. 2007;69:511–34. - PubMed
-
- Hanyaloglu AC, von Zastrow M. Regulation of GPCRs by endocytic membrane trafficking and its potential implications. Annu Rev Pharmacol Toxicol. 2008;48:537–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
