

The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection
- PMID: 21389135
- PMCID: PMC3126191
- DOI: 10.1128/JVI.00198-11
The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection
Abstract
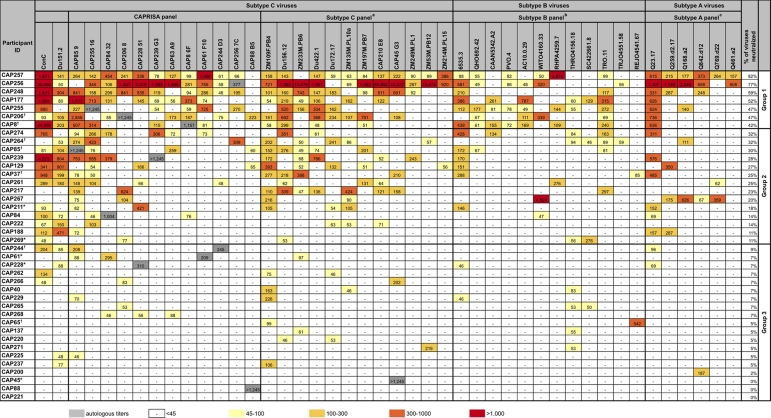
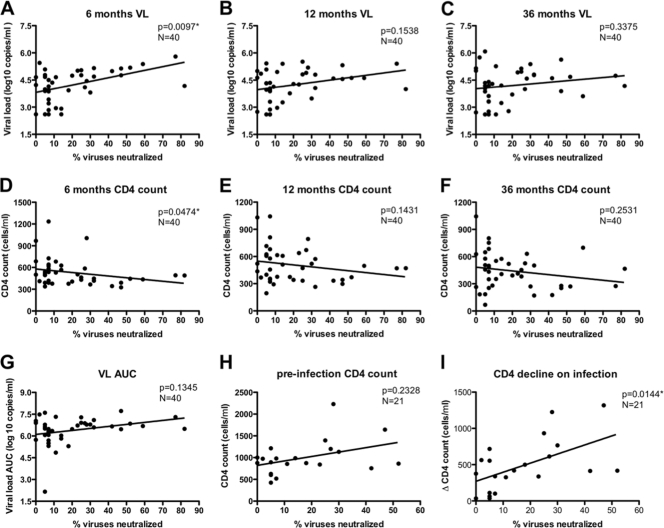
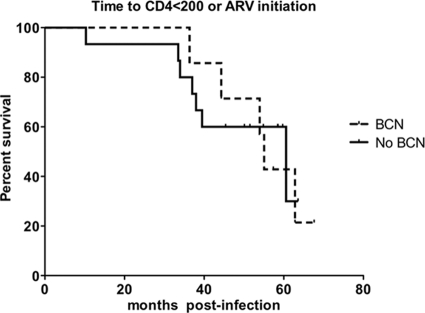
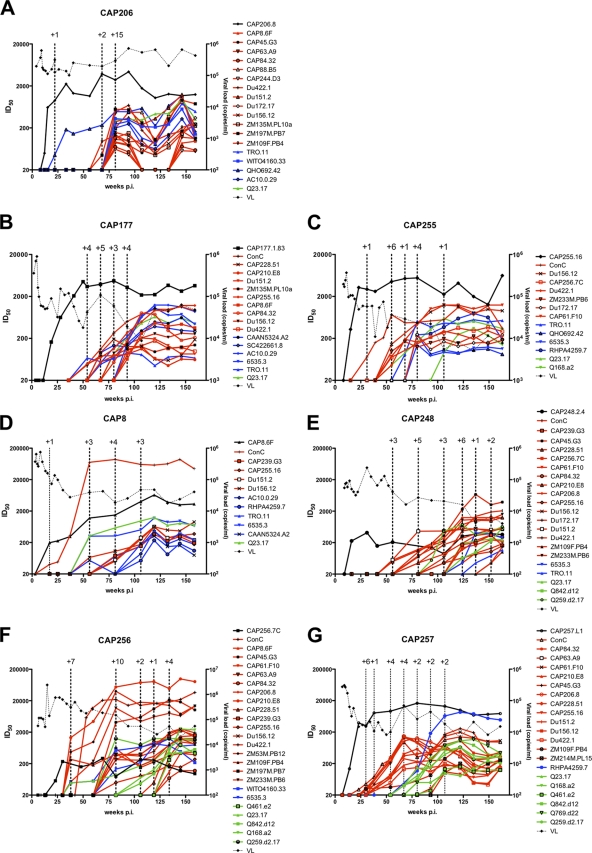
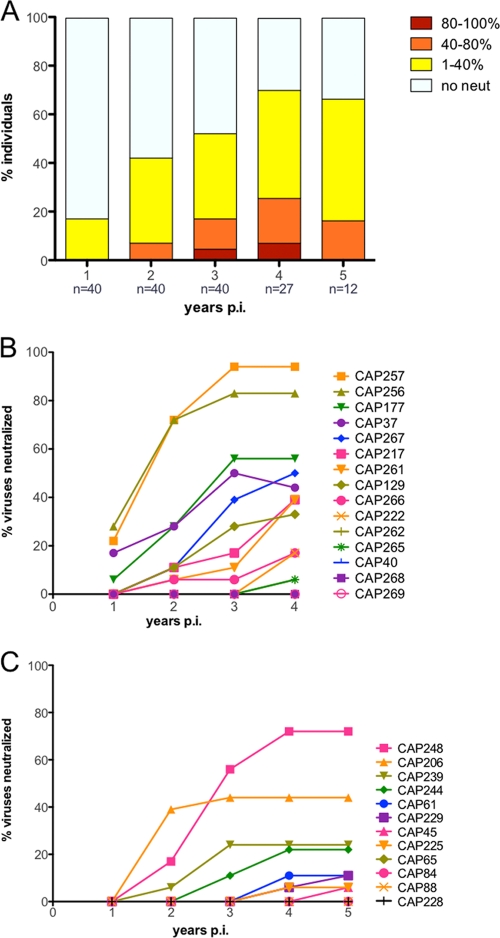
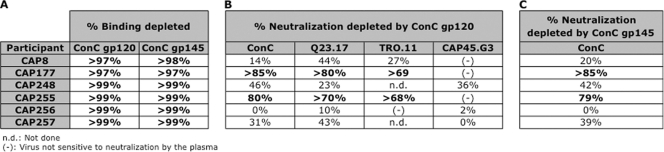
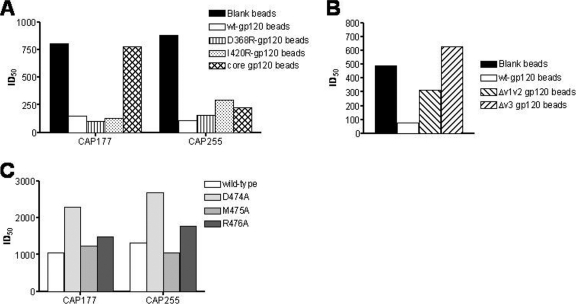
An understanding of how broadly neutralizing activity develops in HIV-1-infected individuals is needed to guide vaccine design and immunization strategies. Here we used a large panel of 44 HIV-1 envelope variants (subtypes A, B, and C) to evaluate the presence of broadly neutralizing antibodies in serum samples obtained 3 years after seroconversion from 40 women enrolled in the CAPRISA 002 acute infection cohort. Seven of 40 participants had serum antibodies that neutralized more than 40% of viruses tested and were considered to have neutralization breadth. Among the samples with breadth, CAP257 serum neutralized 82% (36/44 variants) of the panel, while CAP256 serum neutralized 77% (33/43 variants) of the panel. Analysis of longitudinal samples showed that breadth developed gradually starting from year 2, with the number of viruses neutralized as well as the antibody titer increasing over time. Interestingly, neutralization breadth peaked at 4 years postinfection, with no increase thereafter. The extent of cross-neutralizing activity correlated with CD4(+) T cell decline, viral load, and CD4(+) T cell count at 6 months postinfection but not at later time points, suggesting that early events set the stage for the development of breadth. However, in a multivariate analysis, CD4 decline was the major driver of this association, as viral load was not an independent predictor of breadth. Mapping of the epitopes targeted by cross-neutralizing antibodies revealed that in one individual these antibodies recognized the membrane-proximal external region (MPER), while in two other individuals, cross-neutralizing activity was adsorbed by monomeric gp120 and targeted epitopes that involved the N-linked glycan at position 332 in the C3 region. Serum antibodies from the other four participants targeted quaternary epitopes, at least 2 of which were PG9/16-like and depended on the N160 and/or L165 residue in the V2 region. These data indicate that fewer than 20% of HIV-1 subtype C-infected individuals develop antibodies with cross-neutralizing activity after 3 years of infection and that these antibodies target different regions of the HIV-1 envelope, including as yet uncharacterized epitopes.
Figures
Similar articles
-
Anti-V3/Glycan and Anti-MPER Neutralizing Antibodies, but Not Anti-V2/Glycan Site Antibodies, Are Strongly Associated with Greater Anti-HIV-1 Neutralization Breadth and Potency.J Virol. 2015 May;89(10):5264-75. doi: 10.1128/JVI.00129-15. Epub 2015 Feb 11. J Virol. 2015. PMID: 25673728 Free PMC article.
-
Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop.J Virol. 2011 Apr;85(7):3128-41. doi: 10.1128/JVI.02658-10. Epub 2011 Jan 26. J Virol. 2011. PMID: 21270156 Free PMC article.
-
Viral escape from HIV-1 neutralizing antibodies drives increased plasma neutralization breadth through sequential recognition of multiple epitopes and immunotypes.PLoS Pathog. 2013 Oct;9(10):e1003738. doi: 10.1371/journal.ppat.1003738. Epub 2013 Oct 31. PLoS Pathog. 2013. PMID: 24204277 Free PMC article. Clinical Trial.
-
HIV-1 Subtype C-Infected Children with Exceptional Neutralization Breadth Exhibit Polyclonal Responses Targeting Known Epitopes.J Virol. 2018 Aug 16;92(17):e00878-18. doi: 10.1128/JVI.00878-18. Print 2018 Sep 1. J Virol. 2018. PMID: 29950423 Free PMC article.
-
Engineering strategies of Anti-HIV antibody therapeutics in clinical development.Curr Opin HIV AIDS. 2023 Jul 1;18(4):184-190. doi: 10.1097/COH.0000000000000796. Epub 2023 May 4. Curr Opin HIV AIDS. 2023. PMID: 37144557 Free PMC article. Review.
Cited by
-
Evolution of an HIV glycan-dependent broadly neutralizing antibody epitope through immune escape.Nat Med. 2012 Nov;18(11):1688-92. doi: 10.1038/nm.2985. Epub 2012 Oct 21. Nat Med. 2012. PMID: 23086475 Free PMC article.
-
Progress in HIV-1 vaccine development.Curr Opin HIV AIDS. 2013 Jul;8(4):326-32. doi: 10.1097/COH.0b013e328361d178. Curr Opin HIV AIDS. 2013. PMID: 23743722 Free PMC article. Review.
-
A Bivalent, Chimeric Rabies Virus Expressing Simian Immunodeficiency Virus Envelope Induces Multifunctional Antibody Responses.AIDS Res Hum Retroviruses. 2015 Nov;31(11):1126-38. doi: 10.1089/AID.2014.0319. Epub 2015 May 5. AIDS Res Hum Retroviruses. 2015. PMID: 25848984 Free PMC article.
-
Diverse antibody genetic and recognition properties revealed following HIV-1 envelope glycoprotein immunization.J Immunol. 2015 Jun 15;194(12):5903-14. doi: 10.4049/jimmunol.1500122. Epub 2015 May 11. J Immunol. 2015. PMID: 25964491 Free PMC article.
-
Nonprogressing HIV-infected children share fundamental immunological features of nonpathogenic SIV infection.Sci Transl Med. 2016 Sep 28;8(358):358ra125. doi: 10.1126/scitranslmed.aag1048. Sci Transl Med. 2016. PMID: 27683550 Free PMC article.
References
-
- Baba T. W., et al. 2000. Human neutralizing monoclonal antibodies of the IgG1 subtype protect against mucosal simian-human immunodeficiency virus infection. Nat. Med. 6:200–206 - PubMed
-
- Binley J. 2009. Specificity of broadly neutralizing antibodies in sera from HIV-1 infected individuals. Curr. Opin. HIV AIDS 4:364–372 - PubMed
-
- Blish C. A., Nedellec R., Mandaliya K., Mosier D. E., Overbaugh J. 2007. HIV-1 subtype A envelope variants from early in infection have variable sensitivity to neutralization and to inhibitors of viral entry. AIDS 21:693–702 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
