

Cutaneous immunosurveillance by self-renewing dermal gammadelta T cells
- PMID: 21339323
- PMCID: PMC3058585
- DOI: 10.1084/jem.20101824
Cutaneous immunosurveillance by self-renewing dermal gammadelta T cells
Abstract
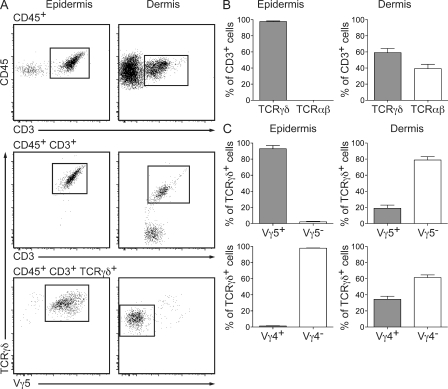
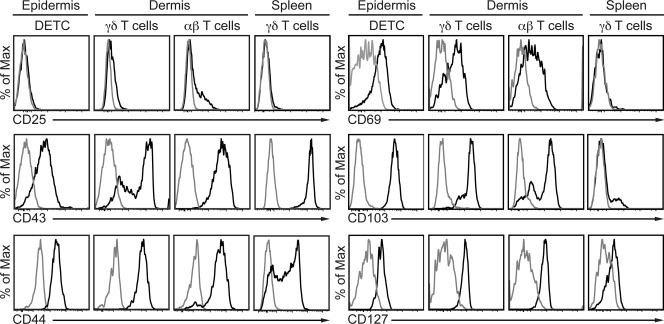
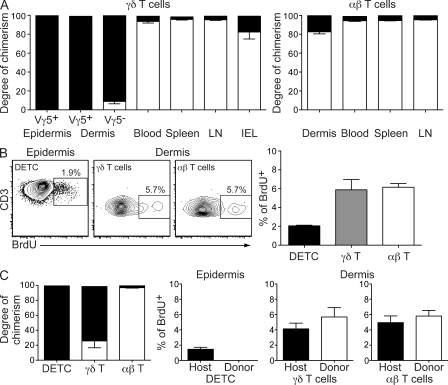
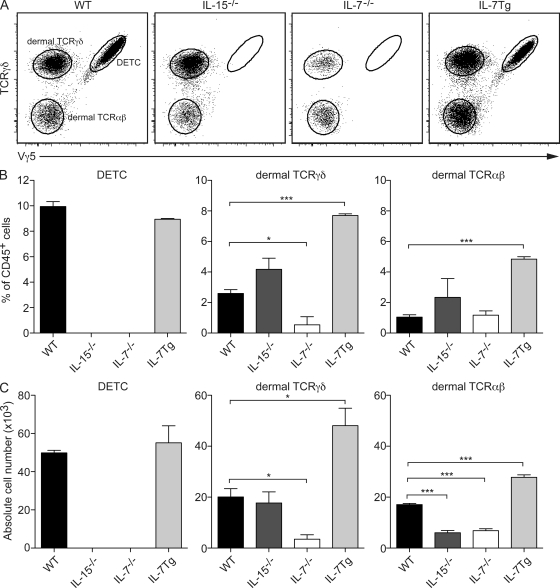
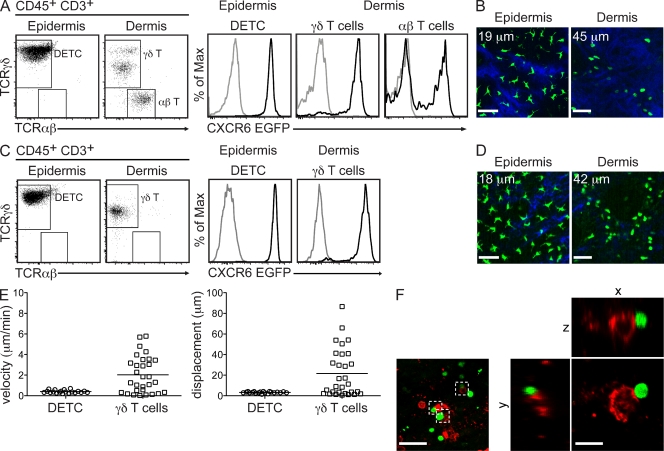
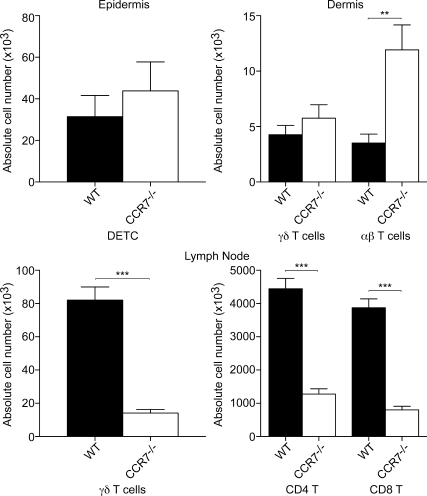
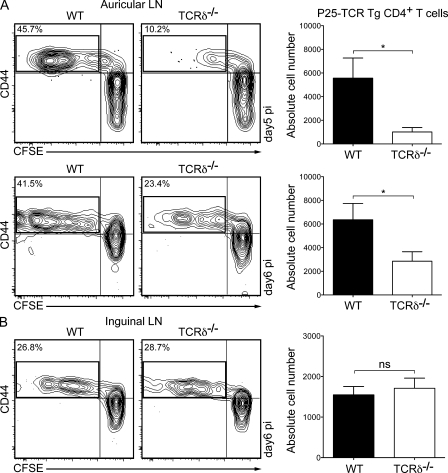
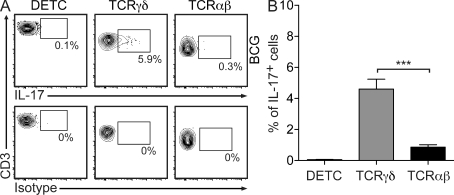
The presence of γδ T cell receptor (TCR)-expressing cells in the epidermis of mice, termed dendritic epidermal T cells (DETCs), is well established. Because of their strict epidermal localization, it is likely that DETCs primarily respond to epithelial stress, such as infections or the presence of transformed cells, whereas they may not participate directly in dermal immune responses. In this study, we describe a prominent population of resident dermal γδ T cells, which differ from DETCs in TCR usage, phenotype, and migratory behavior. Dermal γδ T cells are radioresistant, cycle in situ, and are partially depend on interleukin (IL)-7, but not IL-15, for their development and survival. During mycobacterial infection, dermal γδ T cells are the predominant dermal cells that produce IL-17. Absence of dermal γδ T cells is associated with decreased expansion in skin draining lymph nodes of CD4(+) T cells specific for an immunodominant Mycobacterium tuberculosis epitope. Decreased CD4(+) T cell expansion is related to a reduction in neutrophil recruitment to the skin and decreased BCG shuttling to draining lymph nodes. Thus, dermal γδ T cells are an important part of the resident cutaneous immunosurveillance program. Our data demonstrate functional specialization of T cells in distinct microcompartments of the skin.
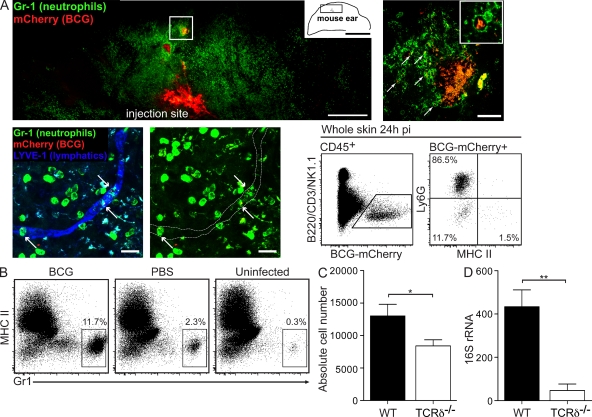
Figures
Similar articles
-
Dermal Vγ4(+) γδ T cells possess a migratory potency to the draining lymph nodes and modulate CD8(+) T-cell activity through TNF-α production.J Invest Dermatol. 2015 Apr;135(4):1007-1015. doi: 10.1038/jid.2014.516. Epub 2014 Dec 10. J Invest Dermatol. 2015. PMID: 25493651
-
IL-1β-dependent activation of dendritic epidermal T cells in contact hypersensitivity.J Immunol. 2014 Apr 1;192(7):2975-83. doi: 10.4049/jimmunol.1301689. Epub 2014 Mar 5. J Immunol. 2014. PMID: 24600030 Free PMC article.
-
Dermal-resident versus recruited γδ T cell response to cutaneous vaccinia virus infection.J Immunol. 2015 Mar 1;194(5):2260-7. doi: 10.4049/jimmunol.1402438. Epub 2015 Jan 21. J Immunol. 2015. PMID: 25609844 Free PMC article.
-
Immunosurveillance and immunoregulation by gammadelta T cells.J Invest Dermatol. 2006 Jan;126(1):25-31. doi: 10.1038/sj.jid.5700003. J Invest Dermatol. 2006. PMID: 16417214 Review.
-
Functions of Vγ4 T Cells and Dendritic Epidermal T Cells on Skin Wound Healing.Front Immunol. 2018 Jun 4;9:1099. doi: 10.3389/fimmu.2018.01099. eCollection 2018. Front Immunol. 2018. PMID: 29915573 Free PMC article. Review.
Cited by
-
Programmed downregulation of CCR6 is important for establishment of epidermal γδT cells by regulating their thymic egress and epidermal location.J Immunol. 2013 Apr 1;190(7):3267-75. doi: 10.4049/jimmunol.1202261. Epub 2013 Feb 18. J Immunol. 2013. PMID: 23420888 Free PMC article.
-
The emerging role of resident memory T cells in protective immunity and inflammatory disease.Nat Med. 2015 Jul;21(7):688-97. doi: 10.1038/nm.3883. Epub 2015 Jun 29. Nat Med. 2015. PMID: 26121195 Free PMC article. Review.
-
The role of chemokines in cutaneous immunosurveillance.Immunol Cell Biol. 2015 Apr;93(4):337-46. doi: 10.1038/icb.2015.16. Epub 2015 Mar 17. Immunol Cell Biol. 2015. PMID: 25776847 Review.
-
Compartmentalized control of skin immunity by resident commensals.Science. 2012 Aug 31;337(6098):1115-9. doi: 10.1126/science.1225152. Epub 2012 Jul 26. Science. 2012. PMID: 22837383 Free PMC article.
-
The Roles of Liver-Resident Lymphocytes in Liver Diseases.Front Immunol. 2019 Jul 16;10:1582. doi: 10.3389/fimmu.2019.01582. eCollection 2019. Front Immunol. 2019. PMID: 31379818 Free PMC article. Review.
References
-
- Abadie V., Badell E., Douillard P., Ensergueix D., Leenen P.J.M., Tanguy M., Fiette L., Saeland S., Gicquel B., Winter N. 2005. Neutrophils rapidly migrate via lymphatics after Mycobacterium bovis BCG intradermal vaccination and shuttle live bacilli to the draining lymph nodes. Blood. 106:1843–1850 10.1182/blood-2005-03-1281 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
