

Structural basis for the recognition and cleavage of histone H3 by cathepsin L
- PMID: 21326229
- PMCID: PMC3105313
- DOI: 10.1038/ncomms1204
Structural basis for the recognition and cleavage of histone H3 by cathepsin L
Abstract
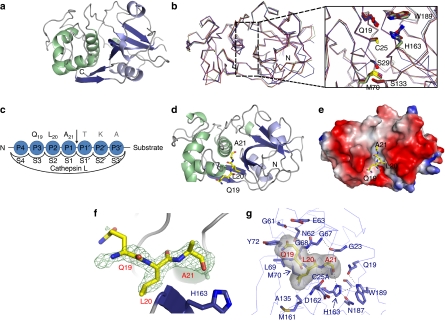
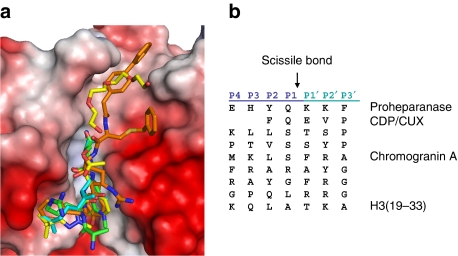
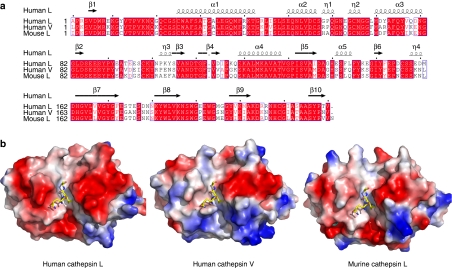
Proteolysis of eukaryotic histone tails has emerged as an important factor in the modulation of cell-cycle progression and cellular differentiation. The recruitment of lysosomal cathepsin L to the nucleus where it mediates proteolysis of the mouse histone H3 tail has been described recently. Here, we report the three-dimensional crystal structures of a mature, inactive mutant of human cathepsin L alone and in complex with a peptide derived from histone H3. Canonical substrate-cathepsin L interactions are observed in the complex between the protease and the histone H3 peptide. Systematic analysis of the impact of posttranslational modifications at histone H3 on substrate selectivity suggests cathepsin L to be highly accommodating of all modified peptides. This is the first report of cathepsin L-histone H3 interaction and the first structural description of cathepsin L in complex with a substrate.
Figures
Similar articles
-
Chasing Tails: Cathepsin-L Improves Structural Analysis of Histones by HX-MS.Mol Cell Proteomics. 2019 Oct;18(10):2089-2098. doi: 10.1074/mcp.RA119.001325. Epub 2019 Aug 13. Mol Cell Proteomics. 2019. PMID: 31409669 Free PMC article.
-
Intestinal differentiation involves cleavage of histone H3 N-terminal tails by multiple proteases.Nucleic Acids Res. 2021 Jan 25;49(2):791-804. doi: 10.1093/nar/gkaa1228. Nucleic Acids Res. 2021. PMID: 33398338 Free PMC article.
-
Cathepsin L proteolytically processes histone H3 during mouse embryonic stem cell differentiation.Cell. 2008 Oct 17;135(2):284-94. doi: 10.1016/j.cell.2008.09.055. Cell. 2008. PMID: 18957203 Free PMC article.
-
Inducible covalent posttranslational modification of histone H3.Sci STKE. 2005 Apr 26;2005(281):re4. doi: 10.1126/stke.2812005re4. Sci STKE. 2005. PMID: 15855410 Review.
-
Cysteine Cathepsins in the secretory vesicle produce active peptides: Cathepsin L generates peptide neurotransmitters and cathepsin B produces beta-amyloid of Alzheimer's disease.Biochim Biophys Acta. 2012 Jan;1824(1):89-104. doi: 10.1016/j.bbapap.2011.08.015. Epub 2011 Sep 8. Biochim Biophys Acta. 2012. PMID: 21925292 Free PMC article. Review.
Cited by
-
DNA and factor VII-activating protease protect against the cytotoxicity of histones.Blood Adv. 2017 Nov 30;1(26):2491-2502. doi: 10.1182/bloodadvances.2017010959. eCollection 2017 Dec 12. Blood Adv. 2017. PMID: 29296900 Free PMC article.
-
Cathepsin X deficiency alters the processing and localisation of cathepsin L and impairs cleavage of a nuclear cathepsin L substrate.Biol Chem. 2024 Feb 28;405(5):351-365. doi: 10.1515/hsz-2023-0355. Print 2024 May 27. Biol Chem. 2024. PMID: 38410910
-
The Roles of Cystatin B in the Brain and Pathophysiological Mechanisms of Progressive Myoclonic Epilepsy Type 1.Cells. 2024 Jan 16;13(2):170. doi: 10.3390/cells13020170. Cells. 2024. PMID: 38247861 Free PMC article. Review.
-
The human anti-HIV antibodies 2F5, 2G12, and PG9 differ in their susceptibility to proteolytic degradation: down-regulation of endogenous serine and cysteine proteinase activities could improve antibody production in plant-based expression platforms.Biotechnol J. 2014 Apr;9(4):493-500. doi: 10.1002/biot.201300207. Biotechnol J. 2014. PMID: 24478053 Free PMC article.
-
Integrative p53, micro-RNA and Cathepsin Protease Co-Regulatory Expression Networks in Cancer.Cancers (Basel). 2020 Nov 20;12(11):3454. doi: 10.3390/cancers12113454. Cancers (Basel). 2020. PMID: 33233599 Free PMC article. Review.
References
-
- Hsing L. C. & Rudensky A. Y. The lysosomal cysteine proteases in MHC class II antigen presentation. Immunol. Rev. 207, 229–241 (2005). - PubMed
-
- Reinheckel T., Deussing J., Roth W. & Peters C. Towards specific functions of lysosomal cysteine peptidases: phenotypes of mice deficient for cathepsin B or cathepsin L. Biol. Chem. 382, 735–741 (2001). - PubMed
-
- Grigoryev S. A., Solovieva V. O., Spirin K. S. & Krasheninnikov I. A. A novel nonhistone protein (MENT) promotes nuclear collapse at the terminal stage of avian erythropoiesis. Exp. Cell Res. 198, 268–275 (1992). - PubMed
-
- Irving J. A. et al.. Inhibitory activity of a heterochromatin-associated serpin (MENT) against papain-like cysteine proteinases affects chromatin structure and blocks cell proliferation. J. Biol. Chem. 277, 13192–13201 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
