

Genome-wide transcription factor binding: beyond direct target regulation
- PMID: 21295369
- PMCID: PMC3068217
- DOI: 10.1016/j.tig.2011.01.001
Genome-wide transcription factor binding: beyond direct target regulation
Abstract
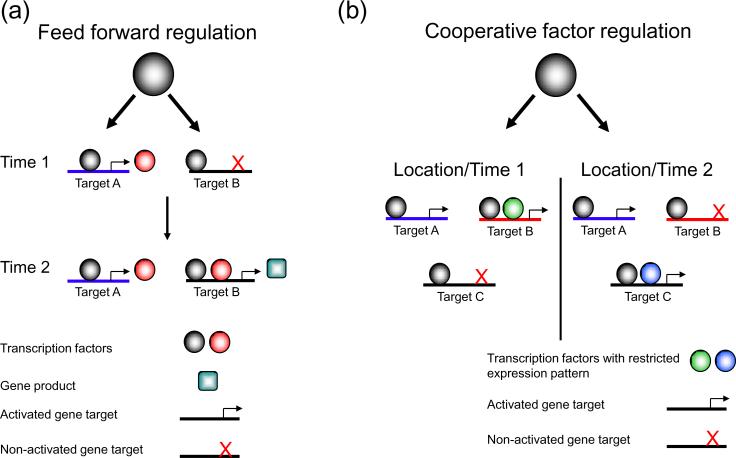
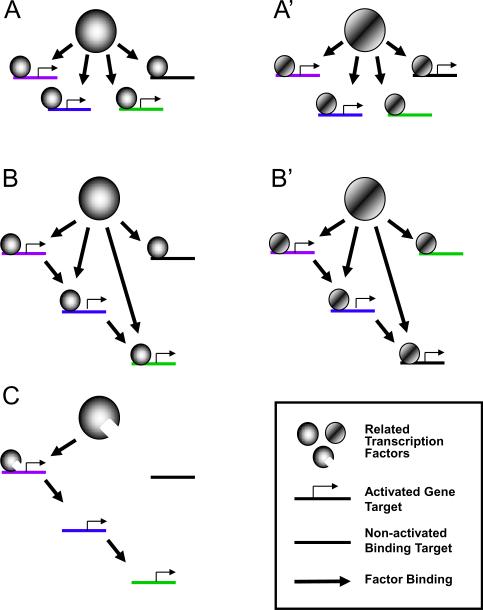
The binding of transcription factors to specific DNA target sequences is the fundamental basis of gene regulatory networks. Chromatin immunoprecipitation combined with DNA tiling arrays or high-throughput sequencing (ChIP-chip and ChIP-seq, respectively) has been used in many recent studies that detail the binding sites of various transcription factors. Surprisingly, data from a variety of model organisms and tissues have demonstrated that transcription factors vary greatly in their number of genomic binding sites, and that binding events can significantly exceed the number of known or possible direct gene targets. Thus, current understanding of transcription factor function must expand to encompass what role, if any, binding might have outside of direct transcriptional target regulation. In this review, we discuss the biological significance of genome-wide binding of transcription factors and present models that can account for this phenomenon.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Integrative analysis of ChIP-chip and ChIP-seq dataset.Methods Mol Biol. 2013;1067:105-24. doi: 10.1007/978-1-62703-607-8_8. Methods Mol Biol. 2013. PMID: 23975789
-
ChIP-Seq Data Analysis to Define Transcriptional Regulatory Networks.Adv Biochem Eng Biotechnol. 2017;160:1-14. doi: 10.1007/10_2016_43. Adv Biochem Eng Biotechnol. 2017. PMID: 28070596 Review.
-
ChIP-ping the branches of the tree: functional genomics and the evolution of eukaryotic gene regulation.Brief Funct Genomics. 2018 Mar 1;17(2):116-137. doi: 10.1093/bfgp/ely004. Brief Funct Genomics. 2018. PMID: 29529131 Free PMC article. Review.
-
Role of ChIP-seq in the discovery of transcription factor binding sites, differential gene regulation mechanism, epigenetic marks and beyond.Cell Cycle. 2014;13(18):2847-52. doi: 10.4161/15384101.2014.949201. Cell Cycle. 2014. PMID: 25486472 Free PMC article. Review.
-
A widespread distribution of genomic CeMyoD binding sites revealed and cross validated by ChIP-Chip and ChIP-Seq techniques.PLoS One. 2010 Dec 29;5(12):e15898. doi: 10.1371/journal.pone.0015898. PLoS One. 2010. PMID: 21209968 Free PMC article.
Cited by
-
Diversity and complexity in chromatin recognition by TFII-I transcription factors in pluripotent embryonic stem cells and embryonic tissues.PLoS One. 2012;7(9):e44443. doi: 10.1371/journal.pone.0044443. Epub 2012 Sep 10. PLoS One. 2012. PMID: 22970219 Free PMC article.
-
Pruning of the adipocyte peroxisome proliferator-activated receptor γ cistrome by hematopoietic master regulator PU.1.Mol Cell Biol. 2013 Aug;33(16):3354-64. doi: 10.1128/MCB.00599-13. Epub 2013 Jun 17. Mol Cell Biol. 2013. PMID: 23775123 Free PMC article.
-
A novel ab initio identification system of transcriptional regulation motifs in genome DNA sequences based on direct comparison scheme of signal/noise distributions.Nucleic Acids Res. 2012 Oct;40(18):8835-48. doi: 10.1093/nar/gks642. Epub 2012 Jul 13. Nucleic Acids Res. 2012. PMID: 22798493 Free PMC article.
-
Disentangling the many layers of eukaryotic transcriptional regulation.Annu Rev Genet. 2012;46:43-68. doi: 10.1146/annurev-genet-110711-155437. Epub 2012 Aug 28. Annu Rev Genet. 2012. PMID: 22934649 Free PMC article. Review.
-
Integrative ChIP-seq/microarray analysis identifies a CTNNB1 target signature enriched in intestinal stem cells and colon cancer.PLoS One. 2014 Mar 20;9(3):e92317. doi: 10.1371/journal.pone.0092317. eCollection 2014. PLoS One. 2014. PMID: 24651522 Free PMC article.
References
-
- Weintraub H. Summary: genetic tinkering--local problems, local solutions. Cold Spring Harb Symp Quant Biol. 1993;58:819–836. - PubMed
-
- Ptashne M, Gann A. Genes & signals. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, New York: 2002. p. xvi.p. 192.
-
- Fry CJ, Peterson CL. Chromatin remodeling enzymes: who's on first? Curr Biol. 2001;11:R185–197. - PubMed
-
- Cosma MP. Ordered recruitment: gene-specific mechanism of transcription activation. Molecular Cell. 2002;10:227–236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
