

Targeting Notch signalling by the conserved miR-8/200 microRNA family in development and cancer cells
- PMID: 21224847
- PMCID: PMC3041954
- DOI: 10.1038/emboj.2010.358
Targeting Notch signalling by the conserved miR-8/200 microRNA family in development and cancer cells
Abstract
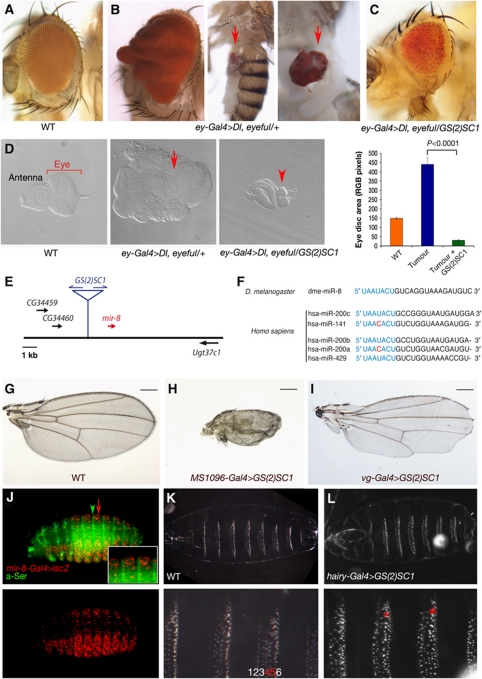
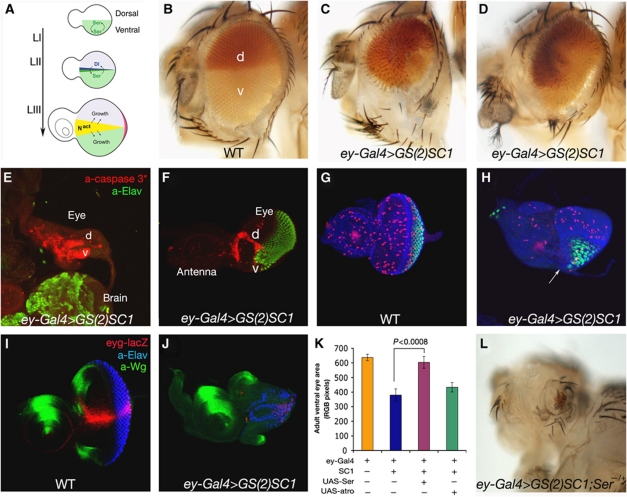
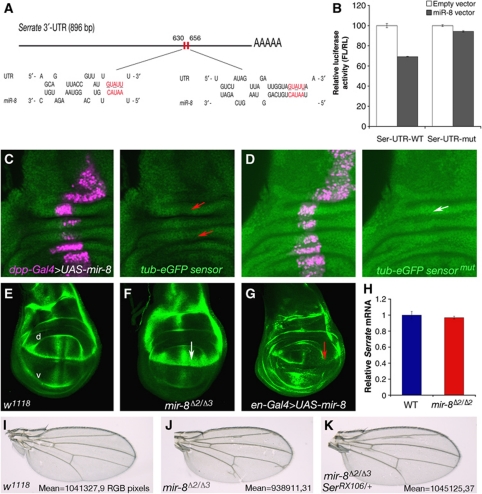
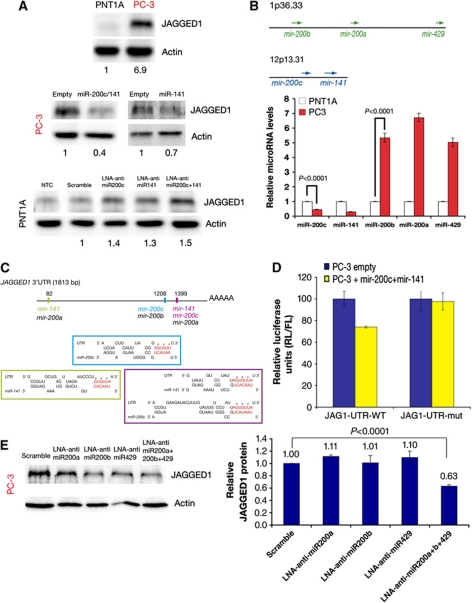
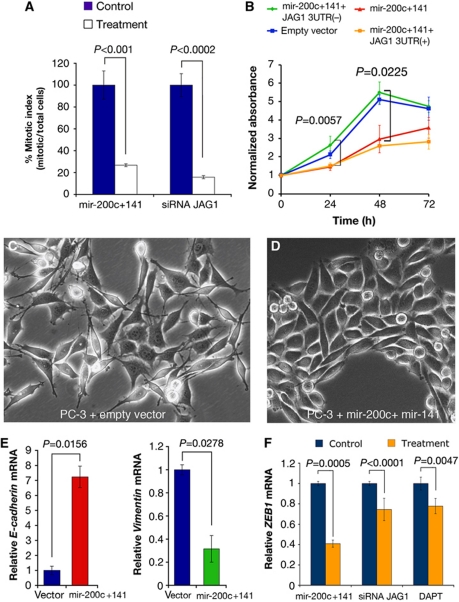
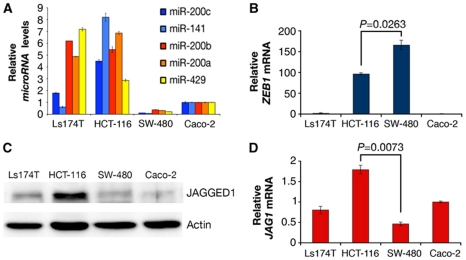
Notch signalling is crucial for the correct development and growth of numerous organs and tissues, and when subverted it can cause cancer. Loss of miR-8/200 microRNAs (miRNAs) is commonly observed in advanced tumours and correlates with their invasion and acquisition of stem-like properties. Here, we show that this miRNA family controls Notch signalling activation in Drosophila and human cells. In an overexpression screen, we identified the Drosophila miR-8 as a potent inhibitor of Notch-induced overgrowth and tumour metastasis. Gain and loss of mir-8 provoked developmental defects reminiscent of impaired Notch signalling and we demonstrated that miR-8 directly inhibits Notch ligand Serrate. Likewise, miR-200c and miR-141 directly inhibited JAGGED1, impeding proliferation of human metastatic prostate cancer cells. It has been suggested that JAGGED1 may also be important for metastases. Although in metastatic cancer cells, JAGGED1 modestly regulated ZEB1, the miR-200c's target in invasion, studies in Drosophila revealed that only concurrent overexpression of Notch and Zfh1/ZEB1 induced tumour metastases. Together, these data define a new way to attenuate or boost Notch signalling that may have clinical interest.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
The ZEB1/miR-200 feedback loop controls Notch signalling in cancer cells.EMBO J. 2011 Feb 16;30(4):770-82. doi: 10.1038/emboj.2010.349. Epub 2011 Jan 11. EMBO J. 2011. PMID: 21224848 Free PMC article.
-
The miR-143/145 cluster is a novel transcriptional target of Jagged-1/Notch signaling in vascular smooth muscle cells.J Biol Chem. 2011 Aug 12;286(32):28312-21. doi: 10.1074/jbc.M111.221945. Epub 2011 Jun 17. J Biol Chem. 2011. PMID: 21685392 Free PMC article.
-
Hypoxia-induced down-regulation of microRNA-34a promotes EMT by targeting the Notch signaling pathway in tubular epithelial cells.PLoS One. 2012;7(2):e30771. doi: 10.1371/journal.pone.0030771. Epub 2012 Feb 17. PLoS One. 2012. PMID: 22363487 Free PMC article.
-
Cell and molecular biology of Notch.J Endocrinol. 2007 Sep;194(3):459-74. doi: 10.1677/JOE-07-0242. J Endocrinol. 2007. PMID: 17761886 Review.
-
Blockade of Jagged/Notch pathway abrogates transforming growth factor β2-induced epithelial-mesenchymal transition in human retinal pigment epithelium cells.Curr Mol Med. 2014 May;14(4):523-34. doi: 10.2174/1566524014666140331230411. Curr Mol Med. 2014. PMID: 24694299 Review.
Cited by
-
Polycomb-mediated silencing of miR-8 is required for maintenance of intestinal stemness in Drosophila melanogaster.Nat Commun. 2024 Mar 2;15(1):1924. doi: 10.1038/s41467-024-46119-9. Nat Commun. 2024. PMID: 38429303 Free PMC article.
-
Nanoparticle-Based RNAi Therapeutics Targeting Cancer Stem Cells: Update and Prospective.Pharmaceutics. 2021 Dec 8;13(12):2116. doi: 10.3390/pharmaceutics13122116. Pharmaceutics. 2021. PMID: 34959397 Free PMC article. Review.
-
Control of microRNA biogenesis and transcription by cell signaling pathways.Curr Opin Genet Dev. 2011 Aug;21(4):504-10. doi: 10.1016/j.gde.2011.04.010. Epub 2011 May 16. Curr Opin Genet Dev. 2011. PMID: 21592778 Free PMC article. Review.
-
Discovery of molecular associations among aging, stem cells, and cancer based on gene expression profiling.Chin J Cancer. 2013 Apr;32(4):155-61. doi: 10.5732/cjc.012.10114. Epub 2013 Jan 9. Chin J Cancer. 2013. PMID: 23298462 Free PMC article. Review.
-
Large-Scale microRNA Expression Profiling Identifies Putative Retinal miRNA-mRNA Signaling Pathways Underlying Form-Deprivation Myopia in Mice.PLoS One. 2016 Sep 13;11(9):e0162541. doi: 10.1371/journal.pone.0162541. eCollection 2016. PLoS One. 2016. PMID: 27622715 Free PMC article.
References
-
- Artavanis-Tsakonas S, Rand MD, Lake RJ (1999) Notch signaling: cell fate control and signal integration in development. Science 284: 770–776 - PubMed
-
- Bailey JM, Singh PK, Hollingsworth MA (2007) Cancer metastasis facilitated by developmental pathways: Sonic hedgehog, Notch, and bone morphogenic proteins. J Cell Biochem 102: 829–839 - PubMed
-
- Bhattacharyya SN, Habermacher R, Martine U, Closs EI, Filipowicz W (2006) Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell 125: 1111–1124 - PubMed
-
- Broihier HT, Moore LA, Van Doren M, Newman S, Lehmann R (1998) zfh-1 is required for germ cell migration and gonadal mesoderm development in Drosophila. Development 125: 655–666 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
