

Progranulin is a chemoattractant for microglia and stimulates their endocytic activity
- PMID: 21224065
- PMCID: PMC3070582
- DOI: 10.1016/j.ajpath.2010.11.002
Progranulin is a chemoattractant for microglia and stimulates their endocytic activity
Abstract
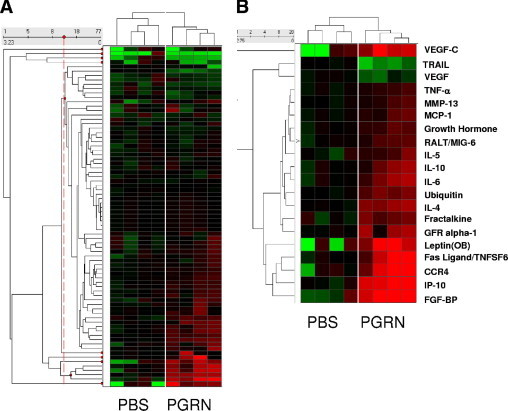
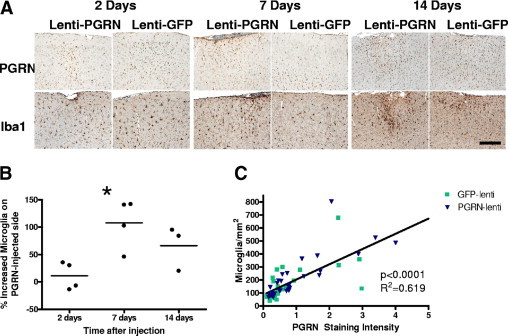
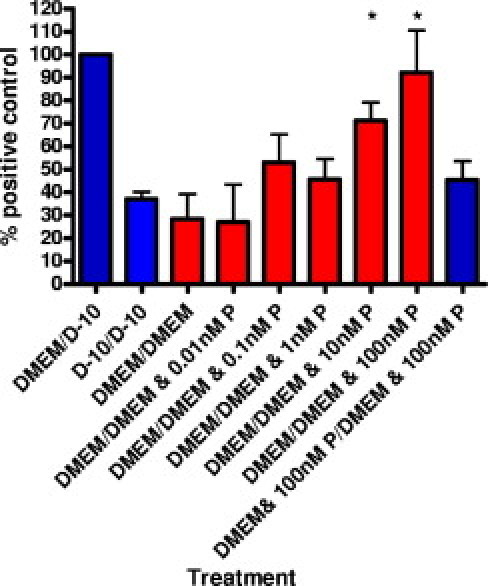
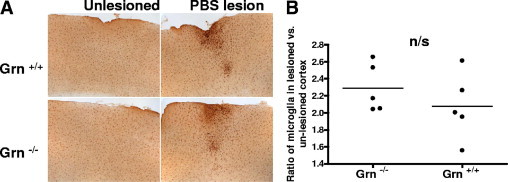
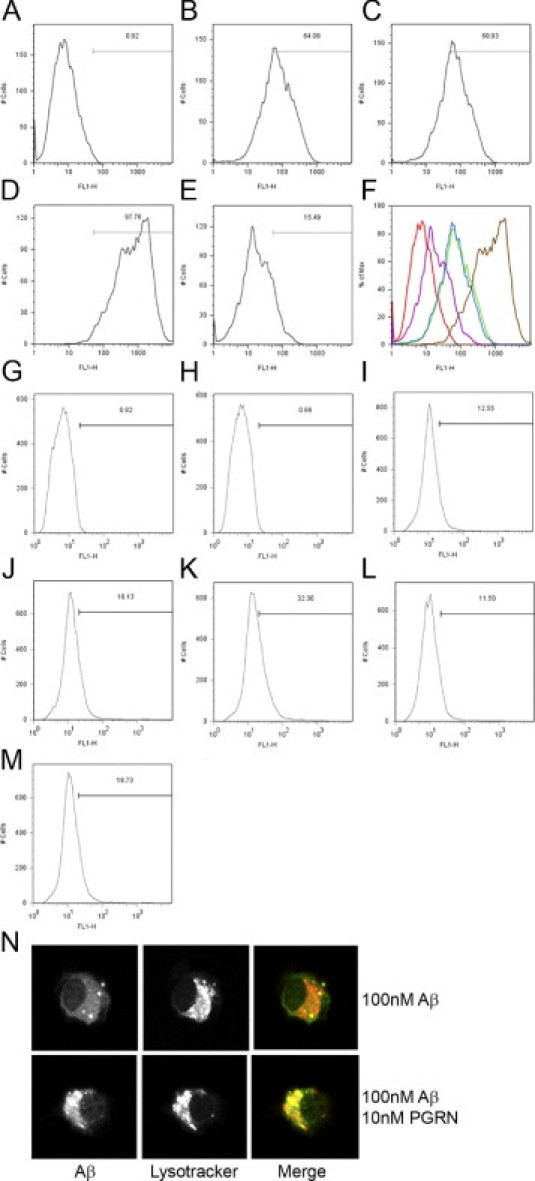
Mutations resulting in progranulin haploinsufficiency cause disease in patients with a subset of frontotemporal lobar degeneration; however, the biological functions of progranulin in the brain remain unknown. To address this subject, the present study initially assessed changes in gene expression and cytokine secretion in rat primary cortical neurons treated with progranulin. Molecular pathways enriched in the progranulin gene set included cell adhesion and cell motility pathways and pathways involved in growth and development. Secretion of cytokines and several chemokines linked to chemoattraction but not inflammation were also increased from progranulin-treated primary neurons. Therefore, whether progranulin is involved in recruitment of immune cells in the brain was investigated. Localized lentiviral expression of progranulin in C57BL/6 mice resulted in an increase of Iba1-positive microglia around the injection site. Moreover, progranulin alone was sufficient to promote migration of primary mouse microglia in vitro. Primary microglia and C4B8 cells demonstrated more endocytosis of amyloid β1-42 when treated with progranulin. These data demonstrate that progranulin acts as a chemoattractant in the brain to recruit or activate microglia and can increase endocytosis of extracellular peptides such as amyloid β.
Copyright © 2011 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Conditional loss of progranulin in neurons is not sufficient to cause neuronal ceroid lipofuscinosis-like neuropathology in mice.Neurobiol Dis. 2017 Oct;106:14-22. doi: 10.1016/j.nbd.2017.06.012. Epub 2017 Jun 21. Neurobiol Dis. 2017. PMID: 28647554
-
Early lysosomal maturation deficits in microglia triggers enhanced lysosomal activity in other brain cells of progranulin knockout mice.Mol Neurodegener. 2018 Sep 4;13(1):48. doi: 10.1186/s13024-018-0281-5. Mol Neurodegener. 2018. PMID: 30180904 Free PMC article.
-
Reduction of microglial progranulin does not exacerbate pathology or behavioral deficits in neuronal progranulin-insufficient mice.Neurobiol Dis. 2019 Apr;124:152-162. doi: 10.1016/j.nbd.2018.11.011. Epub 2018 Nov 15. Neurobiol Dis. 2019. PMID: 30448285 Free PMC article.
-
Cellular effects of progranulin in health and disease.J Mol Neurosci. 2011 Nov;45(3):549-60. doi: 10.1007/s12031-011-9553-z. Epub 2011 May 25. J Mol Neurosci. 2011. PMID: 21611805 Review.
-
Progranulin in frontotemporal lobar degeneration and neuroinflammation.J Neuroinflammation. 2007 Feb 11;4:7. doi: 10.1186/1742-2094-4-7. J Neuroinflammation. 2007. PMID: 17291356 Free PMC article. Review.
Cited by
-
Innate Anti-microbial and Anti-chemotaxis Properties of Progranulin in an Acute Otitis Media Mouse Model.Front Immunol. 2018 Dec 14;9:2952. doi: 10.3389/fimmu.2018.02952. eCollection 2018. Front Immunol. 2018. PMID: 30619312 Free PMC article.
-
TDP-43 and Inflammation: Implications for Amyotrophic Lateral Sclerosis and Frontotemporal Dementia.Int J Mol Sci. 2021 Jul 21;22(15):7781. doi: 10.3390/ijms22157781. Int J Mol Sci. 2021. PMID: 34360544 Free PMC article. Review.
-
Preclinical Interventions in Mouse Models of Frontotemporal Dementia Due to Progranulin Mutations.Neurotherapeutics. 2023 Jan;20(1):140-153. doi: 10.1007/s13311-023-01348-6. Epub 2023 Feb 13. Neurotherapeutics. 2023. PMID: 36781744 Free PMC article. Review.
-
Progranulin inhibits LPS-induced macrophage M1 polarization via NF-кB and MAPK pathways.BMC Immunol. 2020 Jun 5;21(1):32. doi: 10.1186/s12865-020-00355-y. BMC Immunol. 2020. PMID: 32503416 Free PMC article.
-
The established and emerging roles of astrocytes and microglia in amyotrophic lateral sclerosis and frontotemporal dementia.Front Cell Neurosci. 2015 Oct 27;9:414. doi: 10.3389/fncel.2015.00414. eCollection 2015. Front Cell Neurosci. 2015. PMID: 26578880 Free PMC article. Review.
References
-
- Baker M., Mackenzie I.R., Pickering-Brown S.M., Gass J., Rademakers R., Lindholm C., Snowden J., Adamson J., Sadovnick A.D., Rollinson S., Cannon A., Dwosh E., Neary D., Melquist S., Richardson A., Dickson D., Berger Z., Eriksen J., Robinson T., Zehr C., Dickey C.A., Crook R., McGowan E., Mann D., Boeve B., Feldman H., Hutton M. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature. 2006;442:916–919. - PubMed
-
- Rademakers R., Hutton M. The genetics of frontotemporal lobar degeneration. Curr Neurol Neurosci Rep. 2007;7:434–442. - PubMed
-
- Neumann M., Sampathu D.M., Kwong L.K., Truax A.C., Micsenyi M.C., Chou T.T., Bruce J., Schuck T., Grossman M., Clark C.M., McCluskey L.F., Miller B.L., Masliah E., Mackenzie I.R., Feldman H., Feiden W., Kretzschmar H.A., Trojanowski J.Q., Lee V.M.Y. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–133. - PubMed
-
- Brouwers N., Sleegers K., Engelborghs S., Maurer-Stroh S., Gijselinck I., van der Zee J., Pickut B.A., Van den Broeck M., Mattheijssens M., Peeters K., Schymkowitz J., Rousseau F., Martin J.J., Cruts M., De Deyn P.P., Van Broeckhoven C. Genetic variability in progranulin contributes to risk for clinically diagnosed Alzheimer disease. Neurology. 2008;71:656–664. - PubMed
-
- Carecchio M., Fenoglio C., De R.M., Guidi I., Comi C., Cortini F., Venturelli E., Restelli I., Cantoni C., Bresolin N., Monaco F., Scarpini E., Galimberti D. Progranulin plasma levels as potential biomarker for the identification of GRN deletion carriers: a case with atypical onset as clinical amnesic mild cognitive impairment converted to Alzheimer's disease. J Neurol Sci. 2009;287:291–293. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
