

Properties and functions of the nucleocapsid protein in virus assembly
- PMID: 21157181
- PMCID: PMC3073333
- DOI: 10.4161/rna.7.6.14065
Properties and functions of the nucleocapsid protein in virus assembly
Abstract
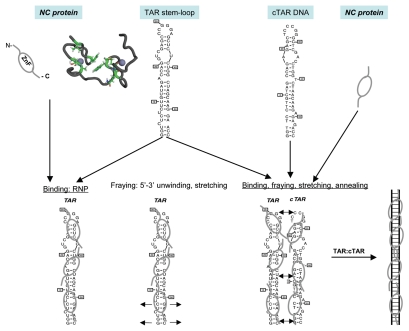
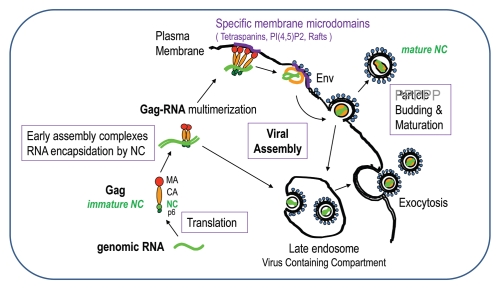
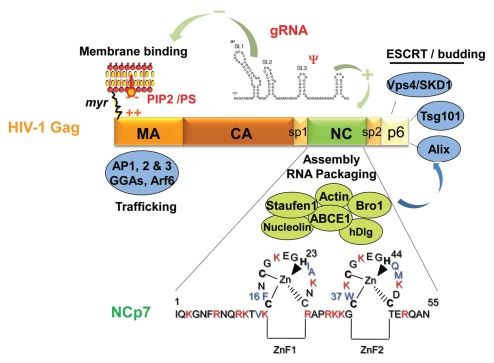
HIV-1 nucleocapsid protein (NC) is a small basic protein generated by the cleavage of the Gag structural polyprotein precusor by the viral protease during virus assembly in the infected cell. HIV-1 NC possesses two copies of a highly conserved CCHC zinc finger (ZnF), flanked by basic residues. HIV-1 NC and more generally retroviral NC proteins are nucleic acid binding proteins possessing potent nucleic acid condensing and chaperoning activities. As such NC protein drives critical structural rearrangements of the genomic RNA, notably RNA dimerization in the course of virus assembly and viral nucleic acid annealing required for genomic RNA replication by the viral reverse transcriptase (RT). Here we review the relationships between the 3D structure of HIV-1 NC, notably the central globular domain encompassing the two zinc fingers and the basic linker and NC functions in the early and late phases of virus replication. One of the salient feature of the NC central globular domain is an hydrophobic plateau which appears to orchestrate the NC functions, such as chaperoning the conversion of the genomic RNA into viral DNA by RT during the early phase, and driving the selection and dimerization of the genomic RNA at the initial stage of viral particle assembly. This ensures a bona fide trafficking of early GagNC-genomic RNA complexes to the plasma membrane of the infected cell and ultimately virion formation and budding.
Figures
Similar articles
-
RNA incorporation is critical for retroviral particle integrity after cell membrane assembly of Gag complexes.J Virol. 2002 Dec;76(23):11853-65. doi: 10.1128/jvi.76.23.11853-11865.2002. J Virol. 2002. PMID: 12414928 Free PMC article.
-
Intracellular HIV-1 Gag localization is impaired by mutations in the nucleocapsid zinc fingers.Retrovirology. 2007 Aug 3;4:54. doi: 10.1186/1742-4690-4-54. Retrovirology. 2007. PMID: 17683545 Free PMC article.
-
Viral RNA annealing activities of human immunodeficiency virus type 1 nucleocapsid protein require only peptide domains outside the zinc fingers.Proc Natl Acad Sci U S A. 1992 Jul 15;89(14):6472-6. doi: 10.1073/pnas.89.14.6472. Proc Natl Acad Sci U S A. 1992. PMID: 1631144 Free PMC article.
-
Features, processing states, and heterologous protein interactions in the modulation of the retroviral nucleocapsid protein function.RNA Biol. 2010 Nov-Dec;7(6):724-34. doi: 10.4161/rna.7.6.13777. Epub 2010 Nov 1. RNA Biol. 2010. PMID: 21045549 Free PMC article. Review.
-
Retroviral Gag protein-RNA interactions: Implications for specific genomic RNA packaging and virion assembly.Semin Cell Dev Biol. 2019 Feb;86:129-139. doi: 10.1016/j.semcdb.2018.03.015. Epub 2018 Apr 1. Semin Cell Dev Biol. 2019. PMID: 29580971 Free PMC article. Review.
Cited by
-
HIV Assembly and Budding: Ca(2+) Signaling and Non-ESCRT Proteins Set the Stage.Mol Biol Int. 2012;2012:851670. doi: 10.1155/2012/851670. Epub 2012 Jun 12. Mol Biol Int. 2012. PMID: 22761998 Free PMC article.
-
When liquid-liquid phase separation meets viral infections.Front Immunol. 2022 Aug 9;13:985622. doi: 10.3389/fimmu.2022.985622. eCollection 2022. Front Immunol. 2022. PMID: 36016945 Free PMC article. Review.
-
The diverse evolutionary histories of domesticated metaviral capsid genes in mammals.bioRxiv [Preprint]. 2023 Sep 17:2023.09.17.558119. doi: 10.1101/2023.09.17.558119. bioRxiv. 2023. Update in: Mol Biol Evol. 2024 Apr 2;41(4):msae061. doi: 10.1093/molbev/msae061 PMID: 37745568 Free PMC article. Updated. Preprint.
-
Recent Advances in HIV-1 Gag Inhibitor Design and Development.Molecules. 2020 Apr 7;25(7):1687. doi: 10.3390/molecules25071687. Molecules. 2020. PMID: 32272714 Free PMC article. Review.
-
RNA Binding Properties of the Ty1 LTR-Retrotransposon Gag Protein.Int J Mol Sci. 2021 Aug 23;22(16):9103. doi: 10.3390/ijms22169103. Int J Mol Sci. 2021. PMID: 34445809 Free PMC article.
References
-
- Baltimore D. RNA-dependent DNA polymerase in virions of RNA tumour viruses. Nature. 1970;226:1209–1211. - PubMed
-
- Gilboa E, Mitra SW, Goff S, Baltimore D. A detailed model of reverse transcription and tests of crucial aspects. Cell. 1979;18:93–100. - PubMed
-
- Coffin JM. In: Virology. Fields BN, Knipe DM, et al., editors. Vol. 2. New York: Raven Press Ltd.; 1990. pp. 1437–500.
-
- Temin HM, Mizutani S. RNA-dependent DNA polymerase in virions of Rous sarcoma virus. Nature. 1970;226:1211–1213. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials