

The C5a receptor impairs IL-12-dependent clearance of Porphyromonas gingivalis and is required for induction of periodontal bone loss
- PMID: 21149611
- PMCID: PMC3075594
- DOI: 10.4049/jimmunol.1003252
The C5a receptor impairs IL-12-dependent clearance of Porphyromonas gingivalis and is required for induction of periodontal bone loss
Abstract
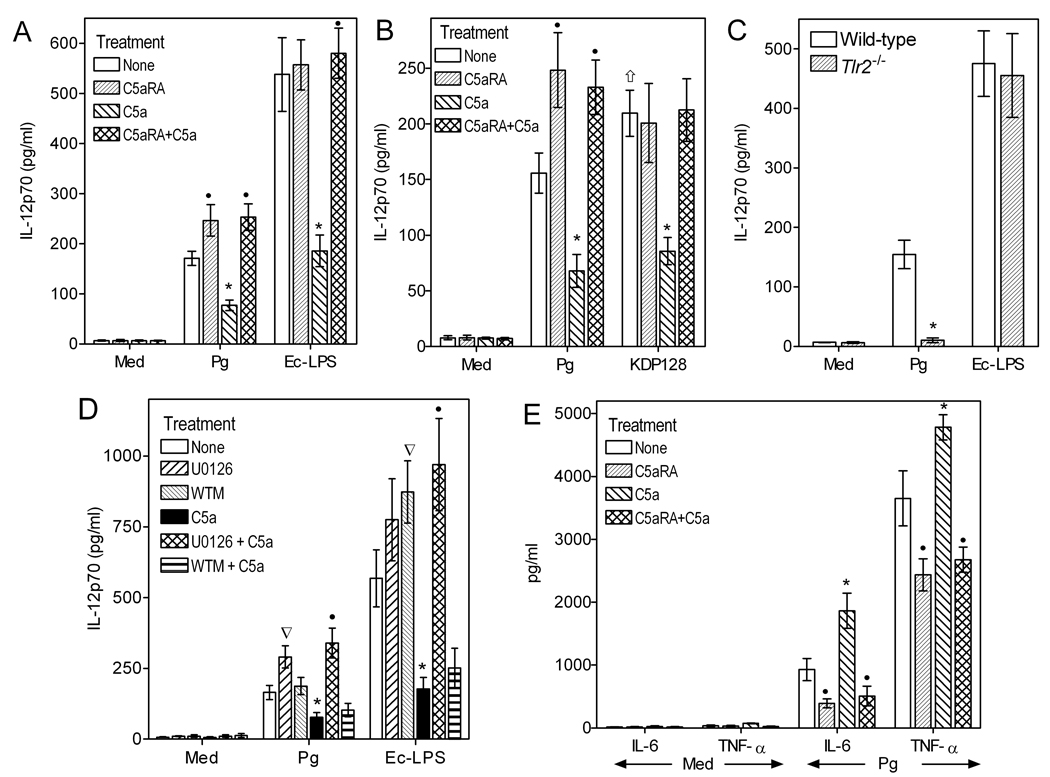
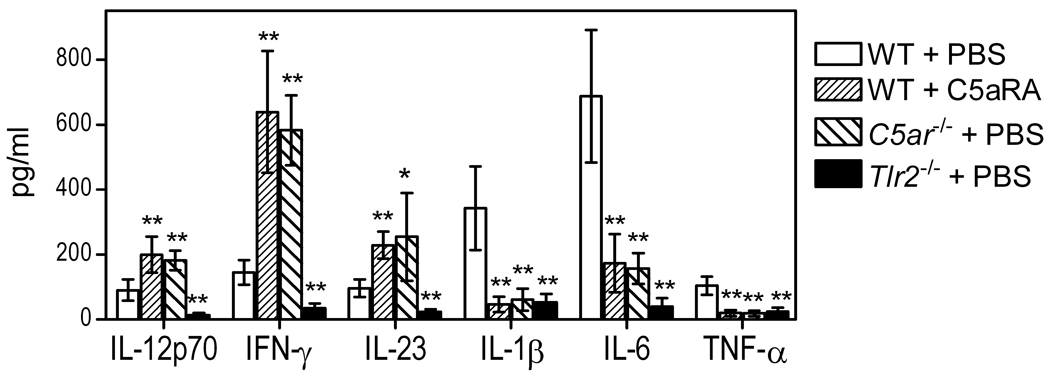
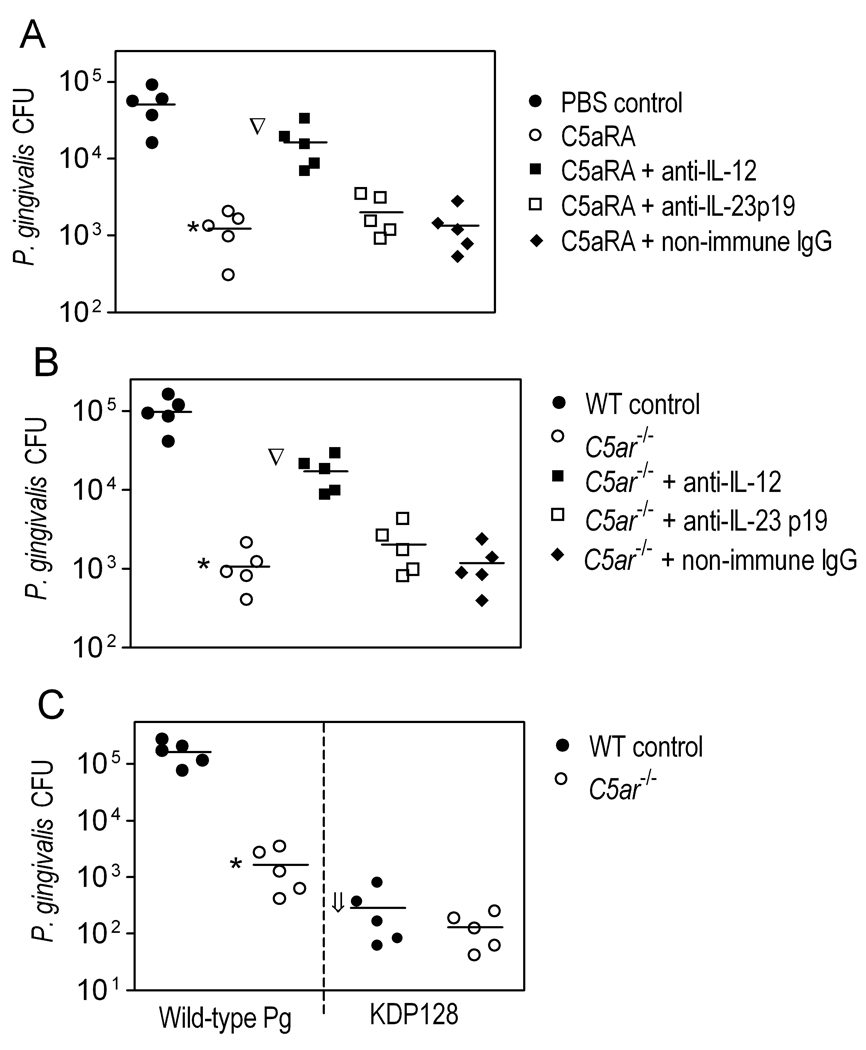
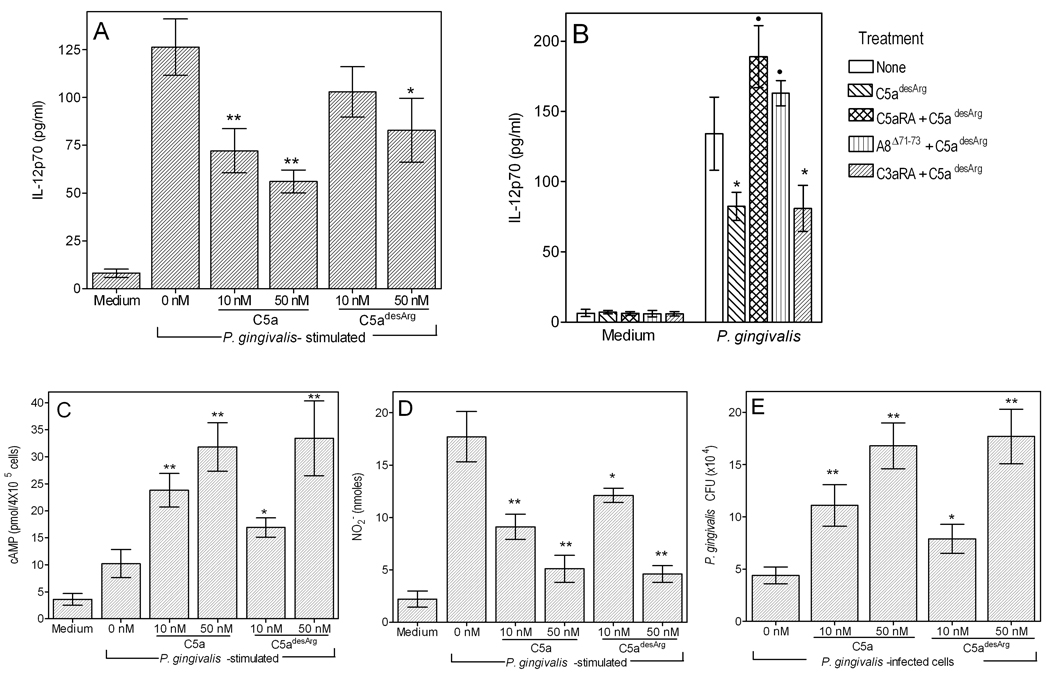
The C5a anaphylatoxin receptor (C5aR; CD88) is activated as part of the complement cascade and exerts important inflammatory, antimicrobial, and regulatory functions, at least in part, via crosstalk with TLRs. However, the periodontal pathogen Porphyromonas gingivalis can control C5aR activation by generating C5a through its own C5 convertase-like enzymatic activity. In this paper, we show that P. gingivalis uses this mechanism to proactively and selectively inhibit TLR2-induced IL-12p70, whereas the same pathogen-instigated C5aR-TLR2 crosstalk upregulates other inflammatory and bone-resorptive cytokines (IL-1β, IL-6, and TNF-α). In vivo, the ability of P. gingivalis to manipulate TLR2 activation via the C5a-C5aR axis allowed it to escape IL-12p70-dependent immune clearance and to cause inflammatory bone loss in a murine model of experimental periodontitis. In the latter regard, C5aR-deficient or TLR2-deficient mice were both resistant to periodontal bone loss, in stark contrast with wild-type control mice, which is consistent with the interdependent interactions of C5aR and TLR2 in P. gingivalis immune evasion and induction of bone-resorptive cytokines. In conclusion, P. gingivalis targets C5aR to promote its adaptive fitness and cause periodontal disease. Given the current availability of safe and effective C5aR antagonists, pharmacological blockade of C5aR could act therapeutically in human periodontitis and reduce associated systemic risks.
Figures
Similar articles
-
Local complement-targeted intervention in periodontitis: proof-of-concept using a C5a receptor (CD88) antagonist.J Immunol. 2012 Dec 1;189(11):5442-8. doi: 10.4049/jimmunol.1202339. Epub 2012 Oct 22. J Immunol. 2012. PMID: 23089394 Free PMC article.
-
Complement receptor 3 blockade promotes IL-12-mediated clearance of Porphyromonas gingivalis and negates its virulence in vivo.J Immunol. 2007 Aug 15;179(4):2359-67. doi: 10.4049/jimmunol.179.4.2359. J Immunol. 2007. PMID: 17675497
-
T cell response mediated by myeloid cell-derived IL-12 is responsible for Porphyromonas gingivalis-induced periodontitis in IL-10-deficient mice.J Immunol. 2008 May 1;180(9):6193-8. doi: 10.4049/jimmunol.180.9.6193. J Immunol. 2008. PMID: 18424741
-
Complement and periodontitis.Biochem Pharmacol. 2010 Dec 15;80(12):1992-2001. doi: 10.1016/j.bcp.2010.06.017. Epub 2010 Jun 23. Biochem Pharmacol. 2010. PMID: 20599785 Free PMC article. Review.
-
The presence, function and regulation of IL-17 and Th17 cells in periodontitis.J Clin Periodontol. 2014 Jun;41(6):541-9. doi: 10.1111/jcpe.12238. Epub 2014 Apr 15. J Clin Periodontol. 2014. PMID: 24735470 Review.
Cited by
-
A novel class of lipoprotein lipase-sensitive molecules mediates Toll-like receptor 2 activation by Porphyromonas gingivalis.Infect Immun. 2013 Apr;81(4):1277-86. doi: 10.1128/IAI.01036-12. Epub 2013 Feb 4. Infect Immun. 2013. PMID: 23381996 Free PMC article.
-
The keystone-pathogen hypothesis.Nat Rev Microbiol. 2012 Oct;10(10):717-25. doi: 10.1038/nrmicro2873. Epub 2012 Sep 3. Nat Rev Microbiol. 2012. PMID: 22941505 Free PMC article. Review.
-
Breaking bad: manipulation of the host response by Porphyromonas gingivalis.Eur J Immunol. 2014 Feb;44(2):328-38. doi: 10.1002/eji.201344202. Eur J Immunol. 2014. PMID: 24338806 Free PMC article. Review.
-
Using next-generation sequencing to detect oral microbiome change following periodontal interventions: A systematic review.Oral Dis. 2021 Jul;27(5):1073-1089. doi: 10.1111/odi.13405. Epub 2020 May 26. Oral Dis. 2021. PMID: 32390250 Free PMC article. Review.
-
The inflammophilic character of the periodontitis-associated microbiota.Mol Oral Microbiol. 2014 Dec;29(6):248-57. doi: 10.1111/omi.12065. Epub 2014 Sep 8. Mol Oral Microbiol. 2014. PMID: 24976068 Free PMC article. Review.
References
-
- Hajishengallis G, Tapping RI, Harokopakis E, Nishiyama S-I, Ratti P, Schifferle RE, Lyle EA, Triantafilou M, Triantafilou K, Yoshimura F. Differential interactions of fimbriae and lipopolysaccharide from Porphyromonas gingivalis with the Toll-like receptor 2-centred pattern recognition apparatus. Cell. Microbiol. 2006;8:1557–1570. - PubMed
-
- Popadiak K, Potempa J, Riesbeck K, Blom AM. Biphasic effect of gingipains from Porphyromonas gingivalis on the human complement system. J. Immunol. 2007;178:7242–7250. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM062134-08/GM/NIGMS NIH HHS/United States
- R01 DE015254/DE/NIDCR NIH HHS/United States
- R01 AI-068730/AI/NIAID NIH HHS/United States
- DE017138/DE/NIDCR NIH HHS/United States
- R01 DE017138-07/DE/NIDCR NIH HHS/United States
- R01 GM-62134/GM/NIGMS NIH HHS/United States
- DE018292/DE/NIDCR NIH HHS/United States
- R01 DE018292-05/DE/NIDCR NIH HHS/United States
- P01 AI068730-05/AI/NIAID NIH HHS/United States
- DE021580/DE/NIDCR NIH HHS/United States
- DE015254/DE/NIDCR NIH HHS/United States
- R01 GM062134/GM/NIGMS NIH HHS/United States
- R01 DE017138/DE/NIDCR NIH HHS/United States
- R01 DE018292/DE/NIDCR NIH HHS/United States
- RC4 DE021580-01/DE/NIDCR NIH HHS/United States
- RC4 DE021580/DE/NIDCR NIH HHS/United States
- P01 AI068730/AI/NIAID NIH HHS/United States
- R01 DE015254-09/DE/NIDCR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous