

Review
doi: 10.1085/jgp.201010508.
3(10) helices in channels and other membrane proteins
Affiliations
- PMID: 21115694
- PMCID: PMC2995148
- DOI: 10.1085/jgp.201010508
Item in Clipboard
Review
3(10) helices in channels and other membrane proteins
J Gen Physiol.
2010 Dec.
No abstract available
Figures
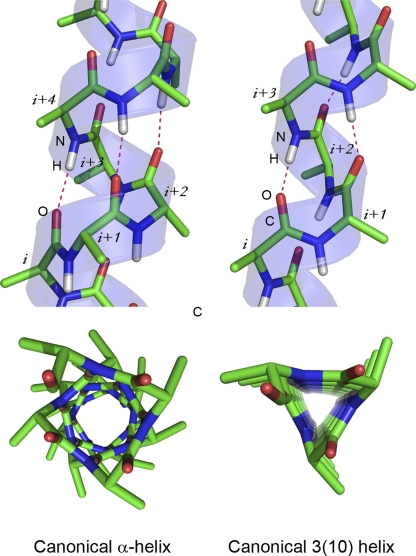
Canonical helical conformations. Side views and views along the axes of a helix in a canonical α-helical conformation (left) and canonical 310 conformation (right). Dotted lines indicate hydrogen bonds between atoms in the backbone of polypeptide. Helices were generated using PepBuild. All figures were prepared with PYMOL.
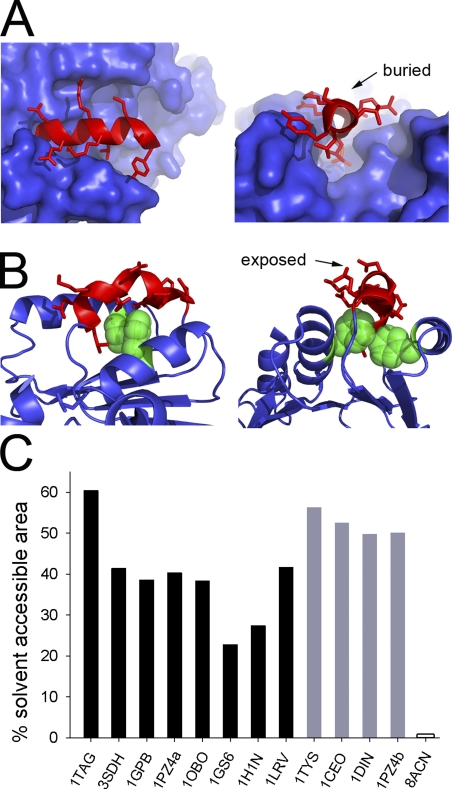
Packing modes of 310 helices. (A) Mode A. Two views of the packing of a 10-residue-long 310 helix (in red) in glycogen phosphorylase (Protein Data Bank accession no. 1GPB). The rest of the protein is shown as blue surface. (B) Mode B. Two views of packing of a 10-residue-long 310 helix (in red) in dienelactone hydrolase (Protein Data Bank accession no. 1DIN). Two phenylalanine residues, shown as green CPK spheres, interact with the face of the 310 helix. Other protein regions are shown as a blue cartoon. (C) Bar graph showing percentage of solvent-accessible surface area for each of the 310 helices identified. Black bars indicate helices in packing mode A. Protein Data Bank accession numbers and residue numbering of respective 310 helices: 1TAG, 201–208; 3SDH, 44–51; 1GPB, 515–524; 1PZ4, 95–103; 1OBO, 1040–1047; 1GS6, 405–412; 1H1N, 180–186; 1LRV, 172–178 (1LRV is a leucine-repeat protein with a 310 helix present in each repeat; we have only considered one repeat in our analysis). Gray bars indicate helices in packing mode B. Protein Data Bank accession numbers and residue numbering of respective 310 helices: 1TYS, 134–140; 1CEO, 241–248; 1DIN, 150–158; 1PZ4, 104–110. Empty bar indicates the fully encased 310 helix (8ACN, 168–174).
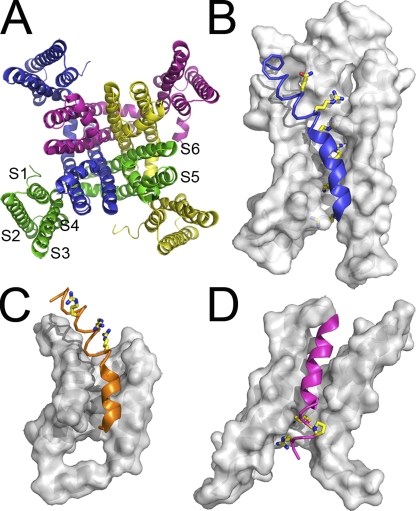
Domain architecture of six-TM channels. (A) Extracellular view of MlotiK1 structure, a bacterial cyclic nucleotide–regulated potassium channel, representative of the superfamily of six-TM helix cation channels. Subunits are shown in different colors. S1 to S4 compose the S1–S4 domain (structurally equivalent to a voltage sensor domain), and S5 and S6 (a pair from each subunit) form the pore domain. The voltage sensor domains of (B) Kv2.1-chimeric voltage-gated potassium channel and of (C) Kv1.2 voltage-gated potassium channel are shown in more detail. (D) The S1–S4 domain of the MlotiK1 channel. Surface representations correspond to S1, S2, and S3. Helices S4 are shown as a thin helical trace along α-helical segments and as a thick ribbon along 310 stretches. Conserved positively charged residues, as well as MlotiK1 proline-108, are shown in stick model. The domains are oriented with the extracellular regions at the top of figure and cytoplasmic regions at the bottom. The N termini of S4 are on the extracellular side of the domains.
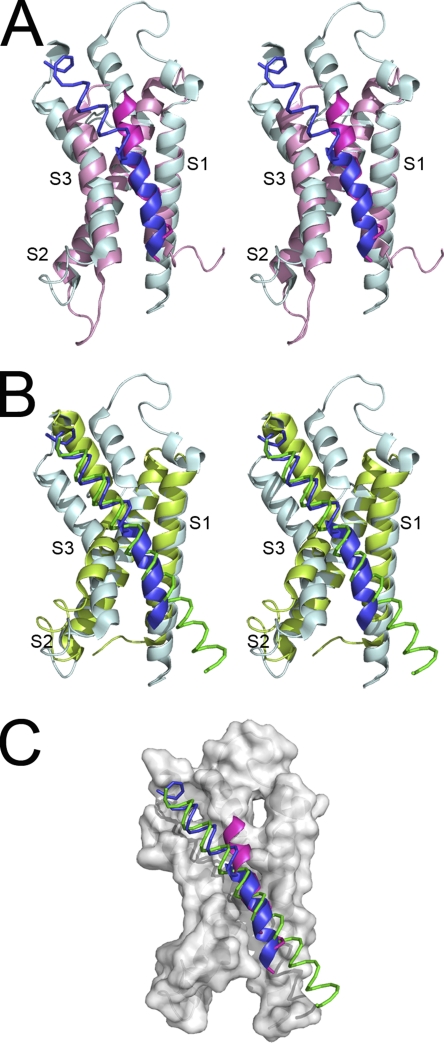
Superposition of voltage sensor and S1–S4 domains. (A) Stereoscopic view of a superposed voltage sensor domain from the Kv2.1-chimeric channel (blue) with the S1–S4 domain from the MlotiK1 channel (magenta). (B) Stereoscopic view of superposed voltage sensor domains from the Kv2.1-chimeric channel (blue) and from the KvAP channel (green). Domains were manually superposed by overlapping S1 and S2 from the different structures. (C) Comparison between the S4 of all three channels using the color code defined in A and B. α-Helical segments are shown as a thin helical trace, and 310 sections are shown as a thick ribbon. Surface representation corresponds to S1, S2, and S3 from the Kv2.1-chimeric channel.
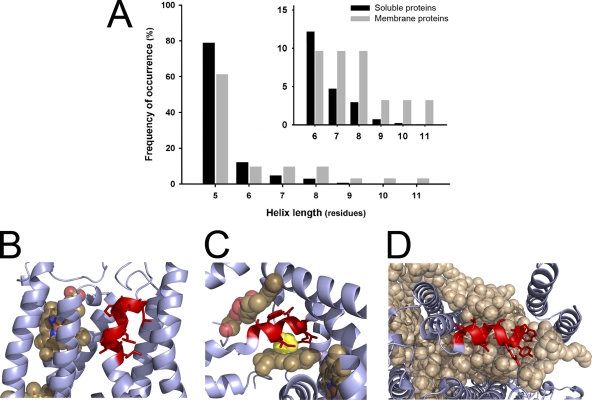
310 helices in other membrane proteins. (A) Graph comparing the frequency of 310 helices in soluble and in membrane proteins. Only 310 helices with lengths between 5 and 11 residues have been considered. The number of helices in each length category was normalized to the total number of detected helices with five or more residues. Inset shows in more detail the differences between membrane and soluble proteins for helices longer than six residues. The data for 310 helices from soluble proteins has been extracted from Enkhbayar et al. (2006). Examples of 310 helices in membrane proteins: 310 helices are shown in red, other protein regions are shown as a blue ribbon, and prosthetic groups are shown as CPK spheres, unless otherwise indicated. (B) 310 helix in cytochrome bc1 complex (Protein Data Bank accession no. 1FYU). Protoporphyrin IX containing Fe (heme) is shown in close proximity to the helix. (C) In bacterial nitrate reductase A (Protein Data Bank accession no. 1Q16), the iron atom from the heme is coordinated by histidine (yellow CPK spheres) from the 310 helix. Opposite the heme and on the top side of the figure, there is phosphatidyl glycerol molecule completing the packing of the 310 helix. (D) In the cyanobacterial photosystem II complex (Protein Data Bank accession no. 3BZ1), a 310 helix is found embedded in a layer formed by LMG (1,2-distearoyl-monogalactosyl-diglyceride), β carotene, and chlorophyll A.
Similar articles
-
Oligomeric structure, dynamics, and orientation of membrane proteins from solid-state NMR.Structure. 2006 Dec;14(12):1731-40. doi: 10.1016/j.str.2006.10.002. Structure. 2006. PMID: 17161364 Review.
-
Membrane simulations: bigger and better?Curr Opin Struct Biol. 2000 Apr;10(2):174-81. doi: 10.1016/s0959-440x(00)00066-x. Curr Opin Struct Biol. 2000. PMID: 10753807 Review.
-
A computational model of the HBK2 potassium channel ion pore.Biochem Biophys Res Commun. 1993 Aug 16;194(3):1117-23. doi: 10.1006/bbrc.1993.1937. Biochem Biophys Res Commun. 1993. PMID: 8352767
-
Proline-induced hinges in transmembrane helices: possible roles in ion channel gating.Proteins. 2001 Aug 1;44(2):63-72. doi: 10.1002/prot.1073. Proteins. 2001. PMID: 11391769
-
Membrane protein folding: beyond the two stage model.FEBS Lett. 2003 Nov 27;555(1):122-5. doi: 10.1016/s0014-5793(03)01106-2. FEBS Lett. 2003. PMID: 14630331 Review.
Cited by
-
Structural basis for ion selectivity in TMEM175 K+ channels.Elife. 2020 Apr 8;9:e53683. doi: 10.7554/eLife.53683. Elife. 2020. PMID: 32267231 Free PMC article.
-
Crystal structures of free and ligand-bound forms of the TetR/AcrR-like regulator SCO3201 from Streptomyces coelicolor suggest a novel allosteric mechanism.FEBS J. 2023 Jan;290(2):521-532. doi: 10.1111/febs.16606. Epub 2022 Sep 2. FEBS J. 2023. PMID: 36017630 Free PMC article.
-
Metal Bridge in S4 Segment Supports Helix Transition in Shaker Channel.Biophys J. 2020 Feb 25;118(4):922-933. doi: 10.1016/j.bpj.2019.08.035. Epub 2019 Sep 5. Biophys J. 2020. PMID: 31635788 Free PMC article.
-
The Role of Lipids in CRAC Channel Function.Biomolecules. 2022 Feb 23;12(3):352. doi: 10.3390/biom12030352. Biomolecules. 2022. PMID: 35327543 Free PMC article. Review.
-
A New Secondary Structure Assignment Algorithm Using Cα Backbone Fragments.Int J Mol Sci. 2016 Mar 11;17(3):333. doi: 10.3390/ijms17030333. Int J Mol Sci. 2016. PMID: 26978354 Free PMC article.
References
-
- Bellanda M., Mammi S., Geremia S., Demitri N., Randaccio L., Broxterman Q.B., Kaptein B., Pengo P., Pasquato L., Scrimin P. 2007. Solvent polarity controls the helical conformation of short peptides rich in Calpha-tetrasubstituted amino acids. Chemistry (Easton). 13:407–416 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
