

Distinct roles of septins in cytokinesis: SEPT9 mediates midbody abscission
- PMID: 21059847
- PMCID: PMC2983063
- DOI: 10.1083/jcb.201006031
Distinct roles of septins in cytokinesis: SEPT9 mediates midbody abscission
Abstract
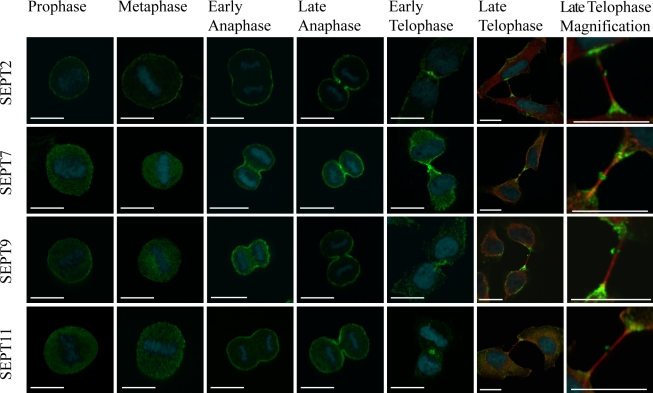
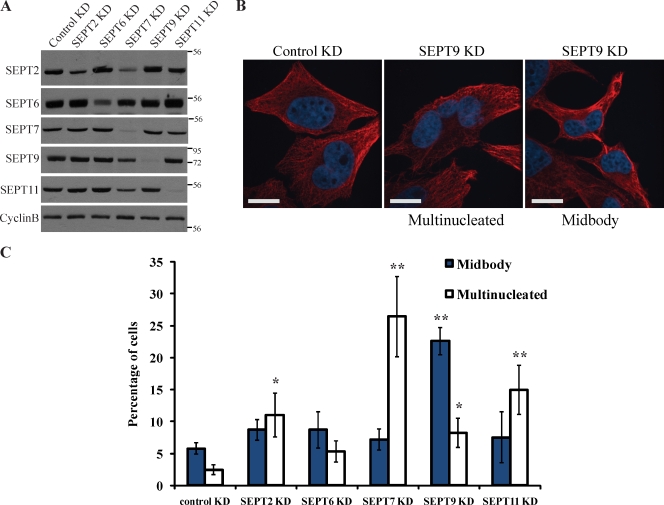
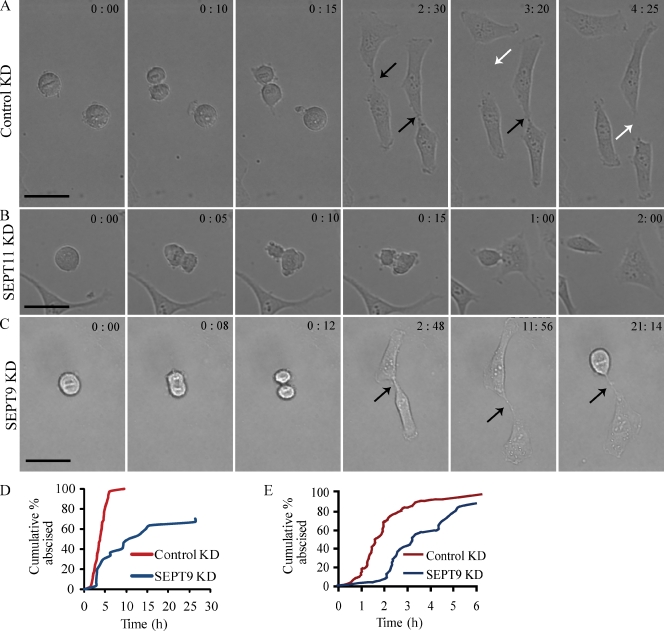
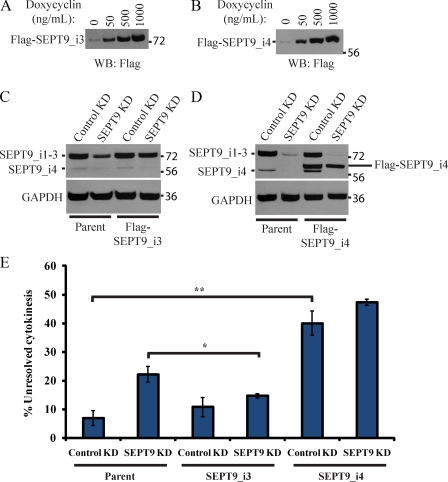
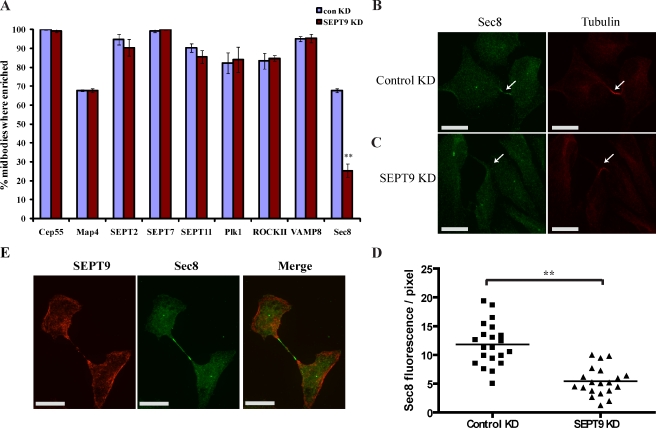
Septins are a family of GTP-binding proteins implicated in mammalian cell division. Most studies examining the role of septins in this process have treated the family as a whole, thus neglecting the possibility that individual members may have diverse functions. To address this, we individually depleted each septin family member expressed in HeLa cells by siRNA and assayed for defects in cell division by immunofluorescence and time-lapse microscopy. Depletion of SEPT2, SEPT7, and SEPT11 causes defects in the early stages of cytokinesis, ultimately resulting in binucleation. In sharp contrast, SEPT9 is dispensable for the early stages of cell division, but is critical for the final separation of daughter cells. Rescue experiments indicate that SEPT9 isoforms containing the N-terminal region are sufficient to drive cytokinesis. We demonstrate that SEPT9 mediates the localization of the vesicle-tethering exocyst complex to the midbody, providing mechanistic insight into the role of SEPT9 during abscission.
Figures
Similar articles
-
SEPT9 occupies the terminal positions in septin octamers and mediates polymerization-dependent functions in abscission.J Cell Biol. 2011 Nov 28;195(5):815-26. doi: 10.1083/jcb.201106131. J Cell Biol. 2011. PMID: 22123865 Free PMC article.
-
Mitotic regulation of SEPT9 protein by cyclin-dependent kinase 1 (Cdk1) and Pin1 protein is important for the completion of cytokinesis.J Biol Chem. 2013 Oct 18;288(42):30075-30086. doi: 10.1074/jbc.M113.474932. Epub 2013 Aug 29. J Biol Chem. 2013. PMID: 23990466 Free PMC article.
-
Cyclophilin A Isomerisation of Septin 2 Mediates Abscission during Cytokinesis.Int J Mol Sci. 2023 Jul 4;24(13):11084. doi: 10.3390/ijms241311084. Int J Mol Sci. 2023. PMID: 37446263 Free PMC article.
-
Sep(t)arate or not – how some cells take septin-independent routes through cytokinesis.J Cell Sci. 2015 May 15;128(10):1877-86. doi: 10.1242/jcs.164830. Epub 2015 Feb 17. J Cell Sci. 2015. PMID: 25690008 Review.
-
The Mammalian Septin Interactome.Front Cell Dev Biol. 2017 Feb 7;5:3. doi: 10.3389/fcell.2017.00003. eCollection 2017. Front Cell Dev Biol. 2017. PMID: 28224124 Free PMC article. Review.
Cited by
-
Steps solidifying a role for SEPT9 in breast cancer suggest that greater strides are needed.Breast Cancer Res. 2012 Jan 9;14(1):101. doi: 10.1186/bcr3056. Breast Cancer Res. 2012. PMID: 22236777 Free PMC article.
-
SEPT9 occupies the terminal positions in septin octamers and mediates polymerization-dependent functions in abscission.J Cell Biol. 2011 Nov 28;195(5):815-26. doi: 10.1083/jcb.201106131. J Cell Biol. 2011. PMID: 22123865 Free PMC article.
-
An efficient protocol for the purification and labeling of entire yeast septin rods from E.coli for quantitative in vitro experimentation.BMC Biotechnol. 2013 Jul 26;13:60. doi: 10.1186/1472-6750-13-60. BMC Biotechnol. 2013. PMID: 23889817 Free PMC article.
-
Tugging at the Heart Strings: The Septin Cytoskeleton in Heart Development and Disease.J Cardiovasc Dev Dis. 2020 Jan 9;7(1):3. doi: 10.3390/jcdd7010003. J Cardiovasc Dev Dis. 2020. PMID: 31936541 Free PMC article. Review.
-
Circulating tumor DNA as an emerging liquid biopsy biomarker for early diagnosis and therapeutic monitoring in hepatocellular carcinoma.Int J Biol Sci. 2020 Mar 5;16(9):1551-1562. doi: 10.7150/ijbs.44024. eCollection 2020. Int J Biol Sci. 2020. PMID: 32226301 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
