

Distinct functions of antigen-specific CD4 T cells during murine Mycobacterium tuberculosis infection
- PMID: 20962277
- PMCID: PMC2984157
- DOI: 10.1073/pnas.1006298107
Distinct functions of antigen-specific CD4 T cells during murine Mycobacterium tuberculosis infection
Abstract
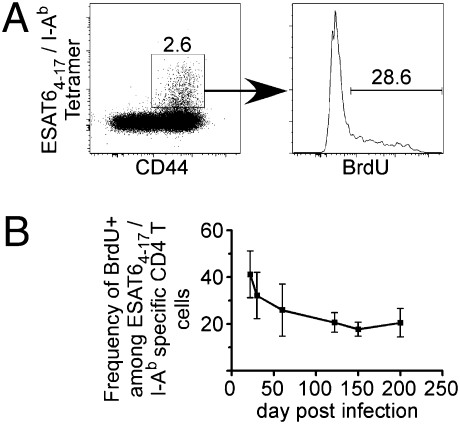
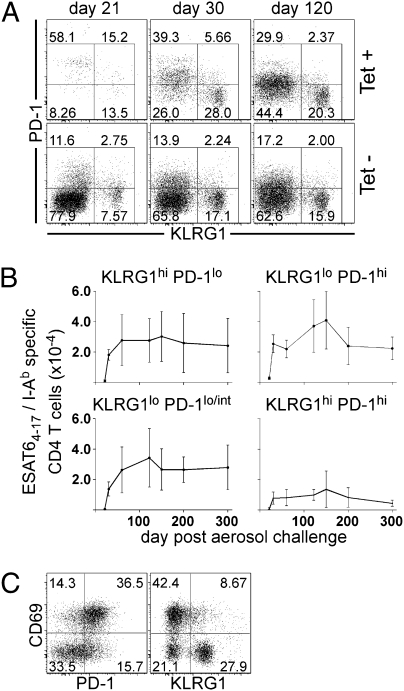
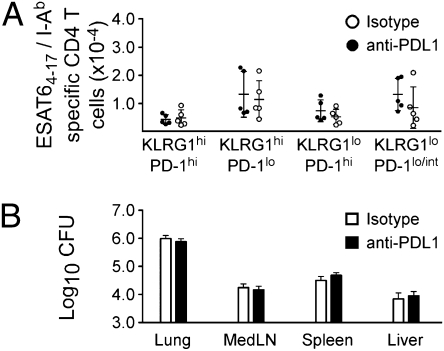
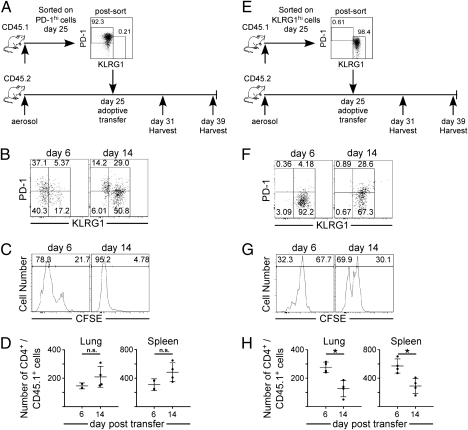
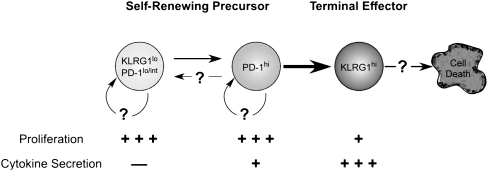
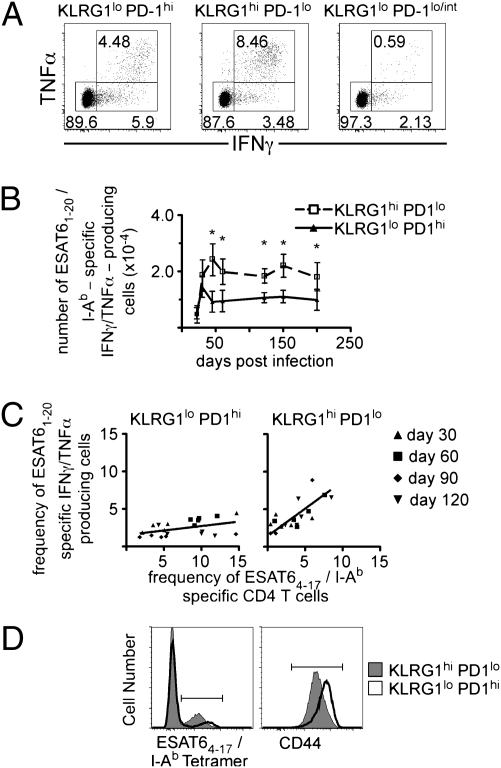
The immune response elicited after Mycobacterium tuberculosis (Mtb) infection is critically dependent on CD4 T cells during both acute and chronic infection. How CD4 T-cell responses are maintained throughout infection is not well understood, and evidence from other infection models has suggested that, under conditions of chronic antigen stimulation, T cells can undergo replicative exhaustion. These findings led us to determine whether subpopulations of CD4 T cells existed that displayed markers of terminal differentiation or exhaustion during murine Mtb infection. Analysis of antigen-specific effector CD4 T cells revealed that programmed death-1 (PD-1) and the killer cell lectin-like receptor G1 (KLRG1) delineated subpopulations of T cells. PD-1-expressing CD4 T cells were highly proliferative, whereas KLRG1 cells exhibited a short lifespan and secreted the cytokines IFNγ and TNFα. Adoptive transfer studies demonstrated that proliferating PD-1-positive CD4 T cells differentiated into cytokine-secreting KLRG1-positive T cells, but not vice versa. Thus, proliferating PD-1-positive cells are not exhausted, but appear to be central to maintaining antigen-specific effector T cells during chronic Mtb infection. Our findings suggest that antigen-specific T-cell responses are maintained during chronic mycobacterial infection through the continual production of terminal effector cells from a proliferating precursor population.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Protective CD4 T cells targeting cryptic epitopes of Mycobacterium tuberculosis resist infection-driven terminal differentiation.J Immunol. 2014 Apr 1;192(7):3247-58. doi: 10.4049/jimmunol.1300283. Epub 2014 Feb 26. J Immunol. 2014. PMID: 24574499
-
Interleukin 27R regulates CD4+ T cell phenotype and impacts protective immunity during Mycobacterium tuberculosis infection.J Exp Med. 2015 Aug 24;212(9):1449-63. doi: 10.1084/jem.20141520. Epub 2015 Aug 17. J Exp Med. 2015. PMID: 26282876 Free PMC article. Clinical Trial.
-
In Vivo Antigen Expression Regulates CD4 T Cell Differentiation and Vaccine Efficacy against Mycobacterium tuberculosis Infection.mBio. 2021 Apr 20;12(2):e00226-21. doi: 10.1128/mBio.00226-21. mBio. 2021. PMID: 33879592 Free PMC article.
-
Defining features of protective CD4 T cell responses to Mycobacterium tuberculosis.Curr Opin Immunol. 2014 Aug;29:137-42. doi: 10.1016/j.coi.2014.06.003. Curr Opin Immunol. 2014. PMID: 25000593 Free PMC article. Review.
-
Physiology and pathology of T-cell aging.Int Immunol. 2020 Apr 12;32(4):223-231. doi: 10.1093/intimm/dxaa006. Int Immunol. 2020. PMID: 31967307 Free PMC article. Review.
Cited by
-
ICOS and Bcl6-dependent pathways maintain a CD4 T cell population with memory-like properties during tuberculosis.J Exp Med. 2015 May 4;212(5):715-28. doi: 10.1084/jem.20141518. Epub 2015 Apr 27. J Exp Med. 2015. PMID: 25918344 Free PMC article.
-
Methamphetamine administration targets multiple immune subsets and induces phenotypic alterations suggestive of immunosuppression.PLoS One. 2012;7(12):e49897. doi: 10.1371/journal.pone.0049897. Epub 2012 Dec 5. PLoS One. 2012. PMID: 23227154 Free PMC article.
-
Therapeutic blockade of PD-L1 and LAG-3 rapidly clears established blood-stage Plasmodium infection.Nat Immunol. 2011 Dec 11;13(2):188-95. doi: 10.1038/ni.2180. Nat Immunol. 2011. PMID: 22157630 Free PMC article.
-
The immunological life cycle of tuberculosis.Nat Rev Immunol. 2012 Jul 13;12(8):581-91. doi: 10.1038/nri3259. Nat Rev Immunol. 2012. PMID: 22790178 Review.
-
Maintenance of peripheral T cell responses during Mycobacterium tuberculosis infection.J Immunol. 2012 Nov 1;189(9):4451-8. doi: 10.4049/jimmunol.1201153. Epub 2012 Oct 1. J Immunol. 2012. PMID: 23028057 Free PMC article.
References
-
- North RJ, Jung YJ. Immunity to tuberculosis. Annu Rev Immunol. 2004;22:599–623. - PubMed
-
- Winslow GM, Roberts AD, Blackman MA, Woodland DL. Persistence and turnover of antigen-specific CD4 T cells during chronic tuberculosis infection in the mouse. J Immunol. 2003;170:2046–2052. - PubMed
-
- Hayflick L. Human cells and aging. Sci Am. 1968;218:32–37. - PubMed
-
- Barber DL, et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439:682–687. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
