

Connexin 43 is critical to maintain the homeostasis of the blood-testis barrier via its effects on tight junction reassembly
- PMID: 20921394
- PMCID: PMC2964254
- DOI: 10.1073/pnas.1007047107
Connexin 43 is critical to maintain the homeostasis of the blood-testis barrier via its effects on tight junction reassembly
Abstract
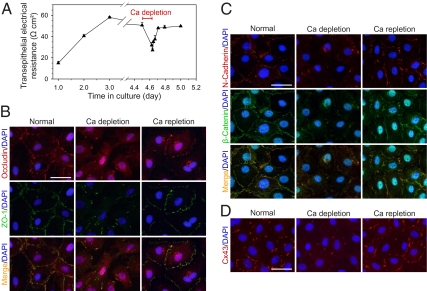
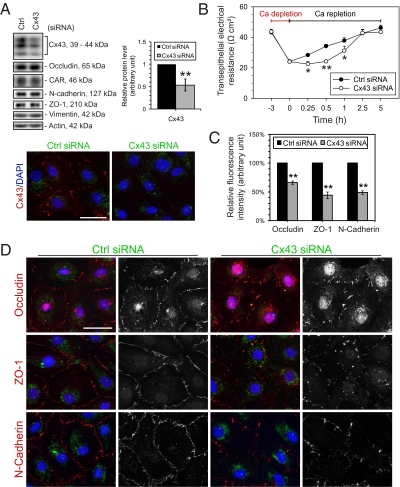
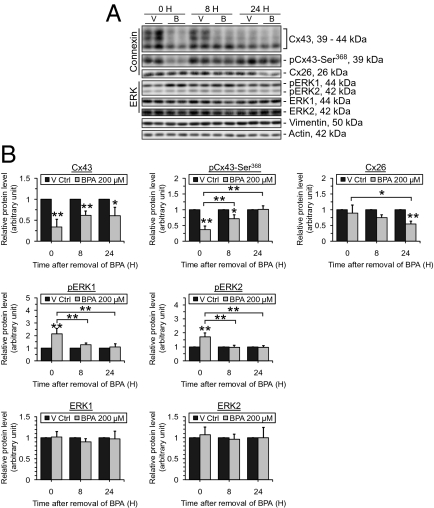
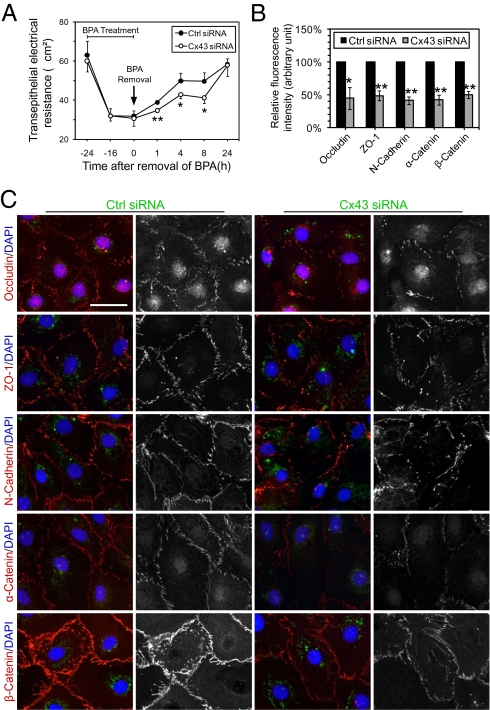
In mammalian testes, the blood-testis barrier (BTB) or Sertoli cell barrier created by specialized junctions between Sertoli cells near the basement membrane confers an immunological barrier by sequestering the events of meiotic division and postmeiotic germ cell development from the systemic circulation. The BTB is constituted by coexisting tight junctions (TJs), basal ectoplasmic specializations, desmosomes, and gap junctions. Despite being one of the tightest blood-tissue barriers, the BTB has to restructure cyclically during spermatogenesis. A recent study showed that gap junction protein connexin 43 (Cx43) and desmosome protein plakophilin-2 are working synergistically to modulate the BTB integrity by regulating the distribution of TJ-associated proteins at the Sertoli-Sertoli cell interface. However, the precise role of Cx43 in regulating the cyclical restructuring of junctions remains obscure. In this report, the calcium switch and the bisphenol A (BPA) models were used to induce junction restructuring in primary cultures of Sertoli cells isolated from rat testes that formed a TJ-permeability barrier that mimicked the BTB in vivo. The removal of calcium by EGTA perturbed the Sertoli cell tight junction barrier, but calcium repletion allowed the "resealing" of the disrupted barrier. However, a knockdown of Cx43 in Sertoli cells by RNAi significantly reduced the kinetics of TJ-barrier resealing. These observations were confirmed using the bisphenol A model in which the knockdown of Cx43 by RNAi also perturbed the TJ-barrier reassembly following BPA removal. In summary, Cx43 is crucial for TJ reassembly at the BTB during its cyclic restructuring throughout the seminiferous epithelial cycle of spermatogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Connexin 43 and plakophilin-2 as a protein complex that regulates blood-testis barrier dynamics.Proc Natl Acad Sci U S A. 2009 Jun 23;106(25):10213-8. doi: 10.1073/pnas.0901700106. Epub 2009 Jun 9. Proc Natl Acad Sci U S A. 2009. PMID: 19509333 Free PMC article.
-
Abcb1a and Abcb1b genes function differentially in blood-testis barrier dynamics in the rat.Cell Death Dis. 2017 Sep 7;8(9):e3038. doi: 10.1038/cddis.2017.435. Cell Death Dis. 2017. PMID: 28880272 Free PMC article.
-
Regulation of blood-testis barrier dynamics by desmosome, gap junction, hemidesmosome and polarity proteins: An unexpected turn of events.Spermatogenesis. 2011 Apr;1(2):105-115. doi: 10.4161/spmg.1.2.15745. Spermatogenesis. 2011. PMID: 22319658 Free PMC article.
-
Cross-talk between tight and anchoring junctions-lesson from the testis.Adv Exp Med Biol. 2008;636:234-54. doi: 10.1007/978-0-387-09597-4_13. Adv Exp Med Biol. 2008. PMID: 19856171 Free PMC article. Review.
-
Extracellular matrix: recent advances on its role in junction dynamics in the seminiferous epithelium during spermatogenesis.Biol Reprod. 2004 Aug;71(2):375-91. doi: 10.1095/biolreprod.104.028225. Epub 2004 Apr 28. Biol Reprod. 2004. PMID: 15115723 Review.
Cited by
-
The Mammalian Blood-Testis Barrier: Its Biology and Regulation.Endocr Rev. 2015 Oct;36(5):564-91. doi: 10.1210/er.2014-1101. Epub 2015 Sep 10. Endocr Rev. 2015. Retraction in: Endocr Rev. 2016 Feb;2016(1):43. doi: 10.1210/er.2015-1127.2016.1.test PMID: 26357922 Free PMC article. Retracted. Review.
-
Rictor/mTORC2 regulates blood-testis barrier dynamics via its effects on gap junction communications and actin filament network.FASEB J. 2013 Mar;27(3):1137-52. doi: 10.1096/fj.12-212977. Epub 2013 Jan 3. FASEB J. 2013. PMID: 23288930 Free PMC article.
-
Lack of the Ig cell adhesion molecule BT-IgSF (IgSF11) induced behavioral changes in the open maze, water maze and resident intruder test.PLoS One. 2023 Jan 6;18(1):e0280133. doi: 10.1371/journal.pone.0280133. eCollection 2023. PLoS One. 2023. PMID: 36607983 Free PMC article.
-
Emerging role for drug transporters at the blood-testis barrier.Trends Pharmacol Sci. 2011 Feb;32(2):99-106. doi: 10.1016/j.tips.2010.11.007. Epub 2010 Dec 17. Trends Pharmacol Sci. 2011. PMID: 21168226 Free PMC article. Review.
-
Mechanisms of environmental chemicals that enable the cancer hallmark of evasion of growth suppression.Carcinogenesis. 2015 Jun;36 Suppl 1(Suppl 1):S2-18. doi: 10.1093/carcin/bgv028. Carcinogenesis. 2015. PMID: 26106139 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
