

Asp3 mediates multiple protein-protein interactions within the accessory Sec system of Streptococcus gordonii
- PMID: 20807195
- PMCID: PMC2959127
- DOI: 10.1111/j.1365-2958.2010.07346.x
Asp3 mediates multiple protein-protein interactions within the accessory Sec system of Streptococcus gordonii
Abstract
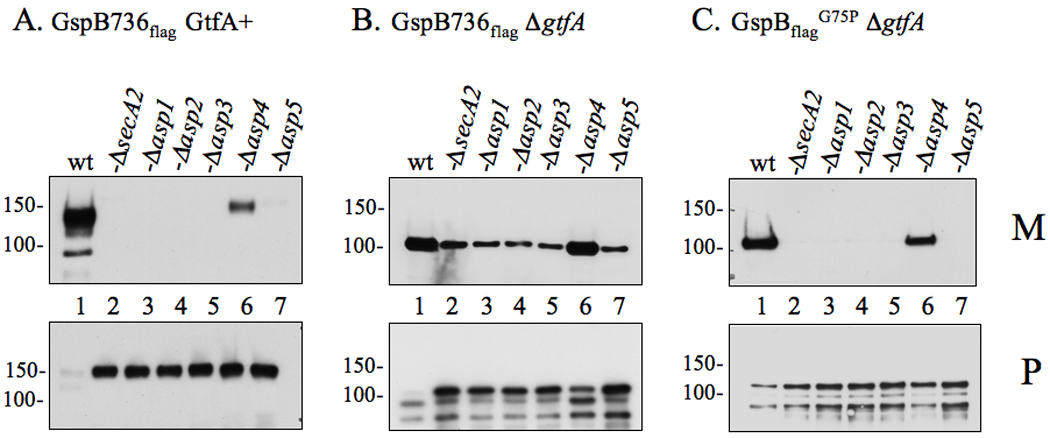
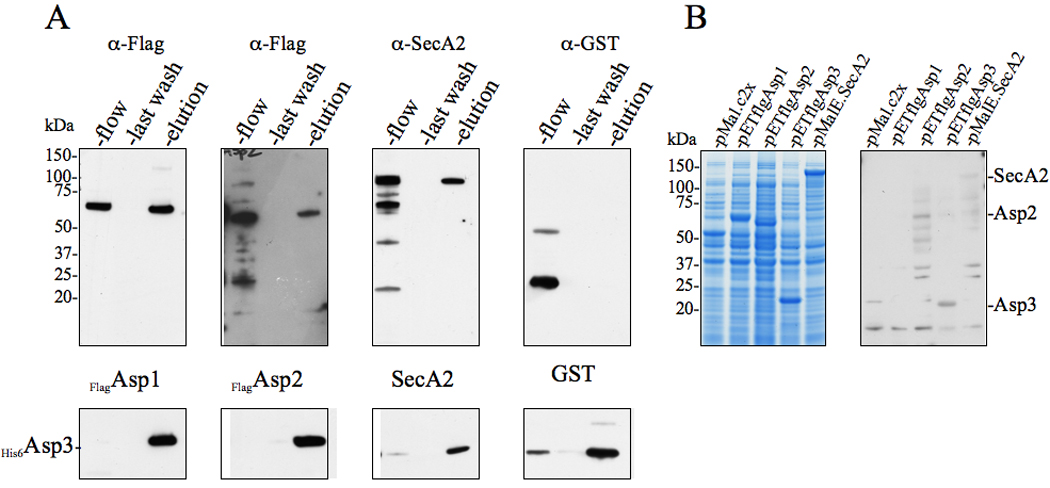
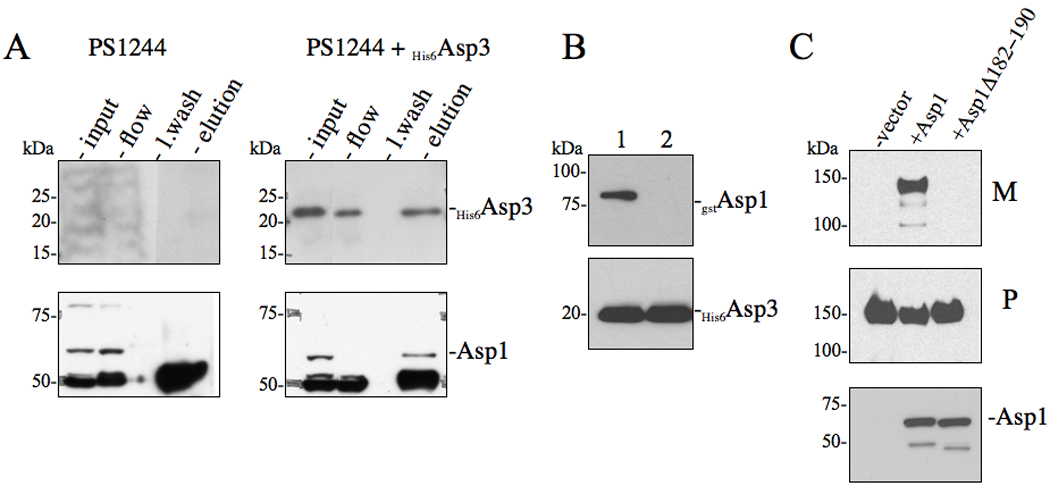
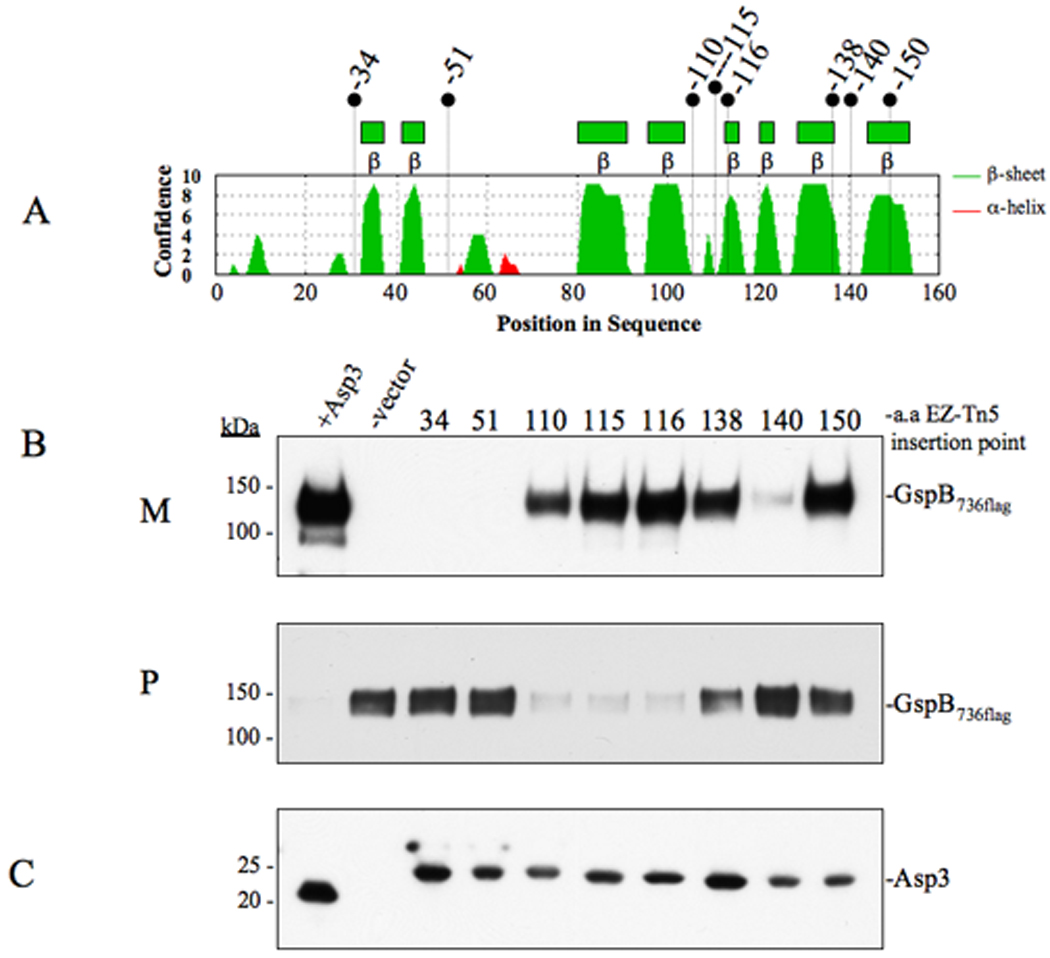
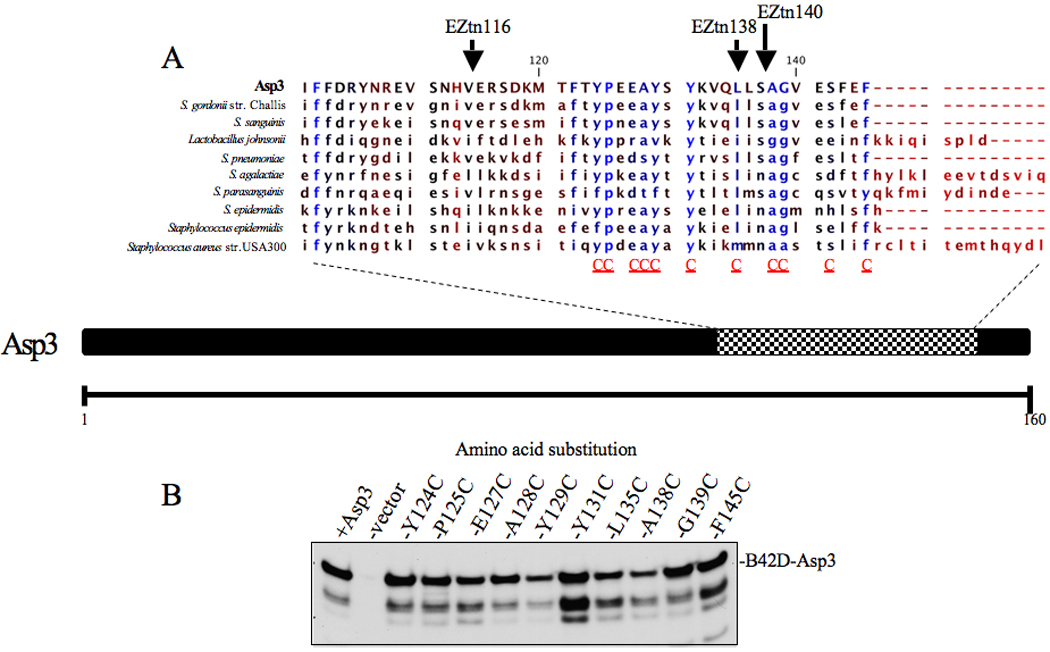
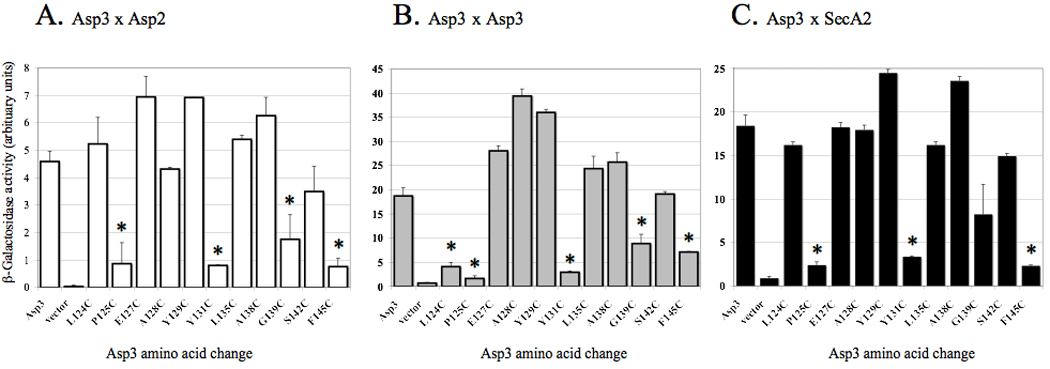
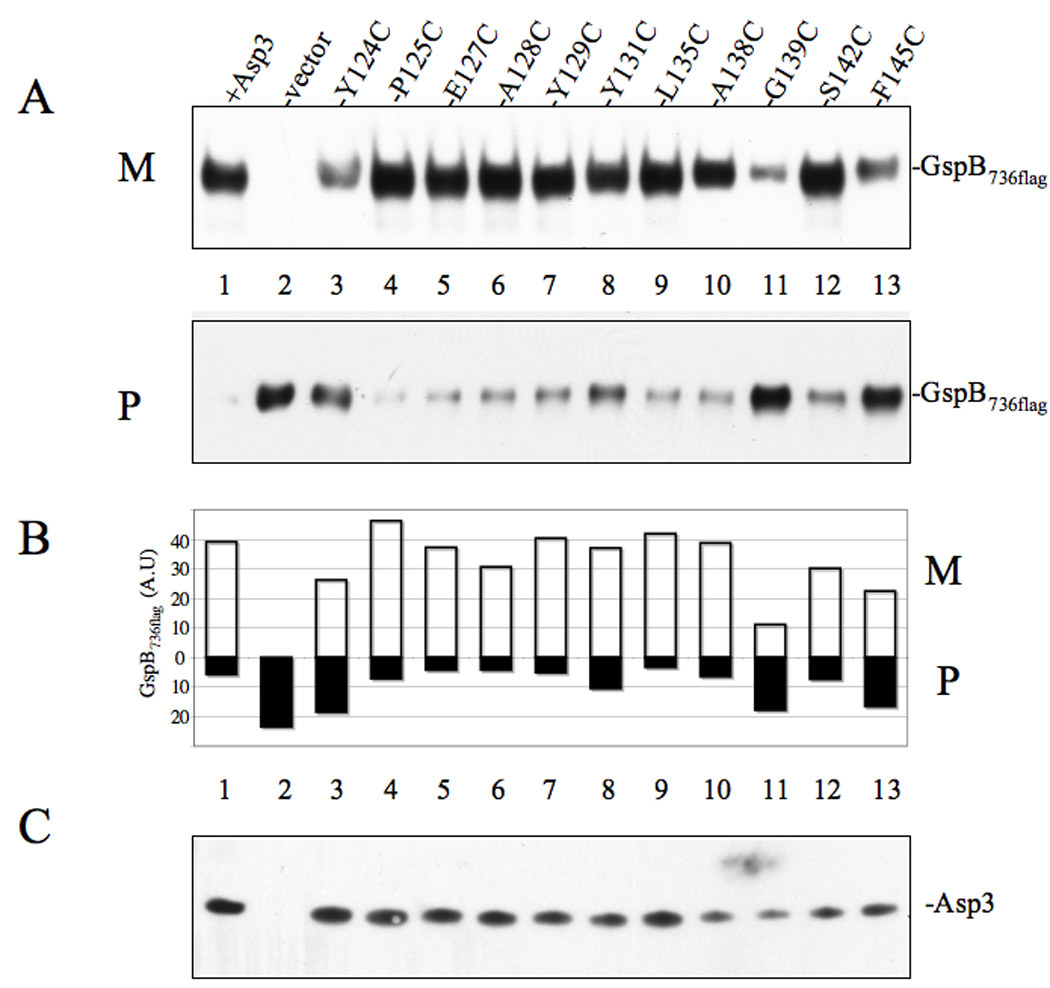
Bacterial binding to human platelets is an important step in the pathogenesis of infective endocarditis. Streptococcus gordonii can mediate its platelet attachment through a cell wall glycoprotein termed GspB ('gordonii surface protein B'). GspB export is mediated by a seven-component accessory Sec system, containing two homologues of the general secretory pathway (SecA2 and SecY2) and five accessory Sec proteins (Asps1-5). Here we show that the Asps are required for optimal export of GspB independent of the glycosylation process. Furthermore, yeast two-hybrid screening of the accessory Sec system revealed interactions occurring between Asp3 and the other components of the system. Asp3 was shown to bind SecA2, Asp1, Asp2 and itself. Mutagenesis of Asp3 identified N- and C-terminal regions that are essential for GspB transport, and conserved residues within the C-terminal domain mediated Asp3 binding to other accessory Sec components. The loss of binding by Asp3 also resulted in an impaired ability of S. gordonii to secrete GspB. These studies indicate that Asp3 is a central element mediating multiple interactions among accessory Sec components that are essential for GspB transport to the cell surface.
Published 2010. This article is a US Government work and is in the public domain in the USA.
Figures

Similar articles
-
Differential localization of the streptococcal accessory sec components and implications for substrate export.J Bacteriol. 2013 Feb;195(4):682-95. doi: 10.1128/JB.01742-12. Epub 2012 Nov 30. J Bacteriol. 2013. PMID: 23204472 Free PMC article.
-
Asp2 and Asp3 interact directly with GspB, the export substrate of the Streptococcus gordonii accessory Sec System.J Bacteriol. 2011 Jul;193(13):3165-74. doi: 10.1128/JB.00057-11. Epub 2011 Apr 29. J Bacteriol. 2011. PMID: 21531800 Free PMC article.
-
Membrane trafficking of the bacterial adhesin GspB and the accessory Sec transport machinery.J Biol Chem. 2019 Feb 1;294(5):1502-1515. doi: 10.1074/jbc.RA118.005657. Epub 2018 Dec 4. J Biol Chem. 2019. PMID: 30514759 Free PMC article.
-
A new twist on an old pathway--accessory Sec [corrected] systems.Mol Microbiol. 2008 Jul;69(2):291-302. doi: 10.1111/j.1365-2958.2008.06294.x. Mol Microbiol. 2008. PMID: 18544071 Free PMC article. Review.
-
The Two Distinct Types of SecA2-Dependent Export Systems.Microbiol Spectr. 2019 May;7(3):10.1128/microbiolspec.psib-0025-2018. doi: 10.1128/microbiolspec.PSIB-0025-2018. Microbiol Spectr. 2019. PMID: 31215505 Free PMC article. Review.
Cited by
-
Differential localization of the streptococcal accessory sec components and implications for substrate export.J Bacteriol. 2013 Feb;195(4):682-95. doi: 10.1128/JB.01742-12. Epub 2012 Nov 30. J Bacteriol. 2013. PMID: 23204472 Free PMC article.
-
Canonical SecA associates with an accessory secretory protein complex involved in biogenesis of a streptococcal serine-rich repeat glycoprotein.J Bacteriol. 2011 Dec;193(23):6560-6. doi: 10.1128/JB.05668-11. Epub 2011 Sep 30. J Bacteriol. 2011. PMID: 21965576 Free PMC article.
-
O-acetylation controls the glycosylation of bacterial serine-rich repeat glycoproteins.J Biol Chem. 2021 Jan-Jun;296:100249. doi: 10.1074/jbc.RA120.016116. Epub 2021 Jan 9. J Biol Chem. 2021. PMID: 33384382 Free PMC article.
-
A Specific interaction between SecA2 and a region of the preprotein adjacent to the signal peptide occurs during transport via the accessory Sec system.J Biol Chem. 2012 Jul 13;287(29):24438-47. doi: 10.1074/jbc.M112.378059. Epub 2012 May 31. J Biol Chem. 2012. PMID: 22654116 Free PMC article.
-
Streptococcal Serine-Rich Repeat Proteins in Colonization and Disease.Front Microbiol. 2020 Oct 30;11:593356. doi: 10.3389/fmicb.2020.593356. eCollection 2020. Front Microbiol. 2020. PMID: 33193266 Free PMC article. Review.
References
-
- Bensing BA, Sullam PM. An accessory sec locus of Streptococcus gordonii is required for export of the surface protein GspB and for normal levels of binding to human platelets. Mol Microbiol. 2002;44:1081–1094. - PubMed
-
- Bensing BA, Takamatsu D, Sullam PM. Determinants of the streptococcal surface glycoprotein GspB that facilitate export by the accessory Sec system. Mol Microbiol. 2005;58:1468–1481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
