

Catalysis of lysine 48-specific ubiquitin chain assembly by residues in E2 and ubiquitin
- PMID: 20797627
- PMCID: PMC2929935
- DOI: 10.1016/j.molcel.2010.07.027
Catalysis of lysine 48-specific ubiquitin chain assembly by residues in E2 and ubiquitin
Abstract
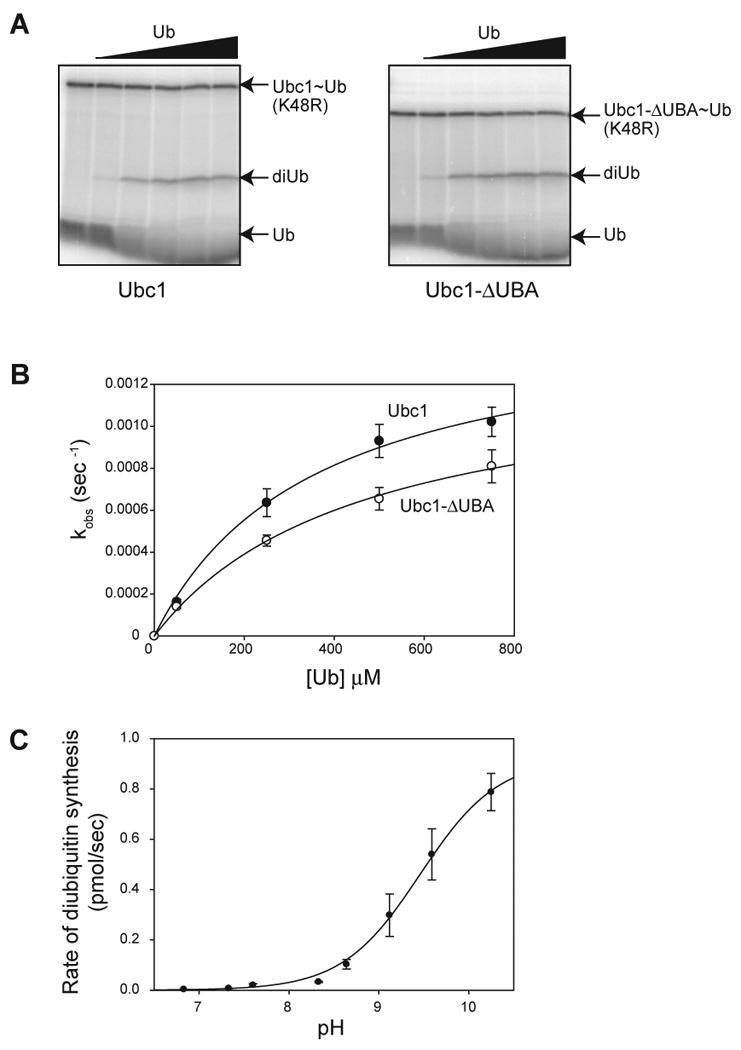
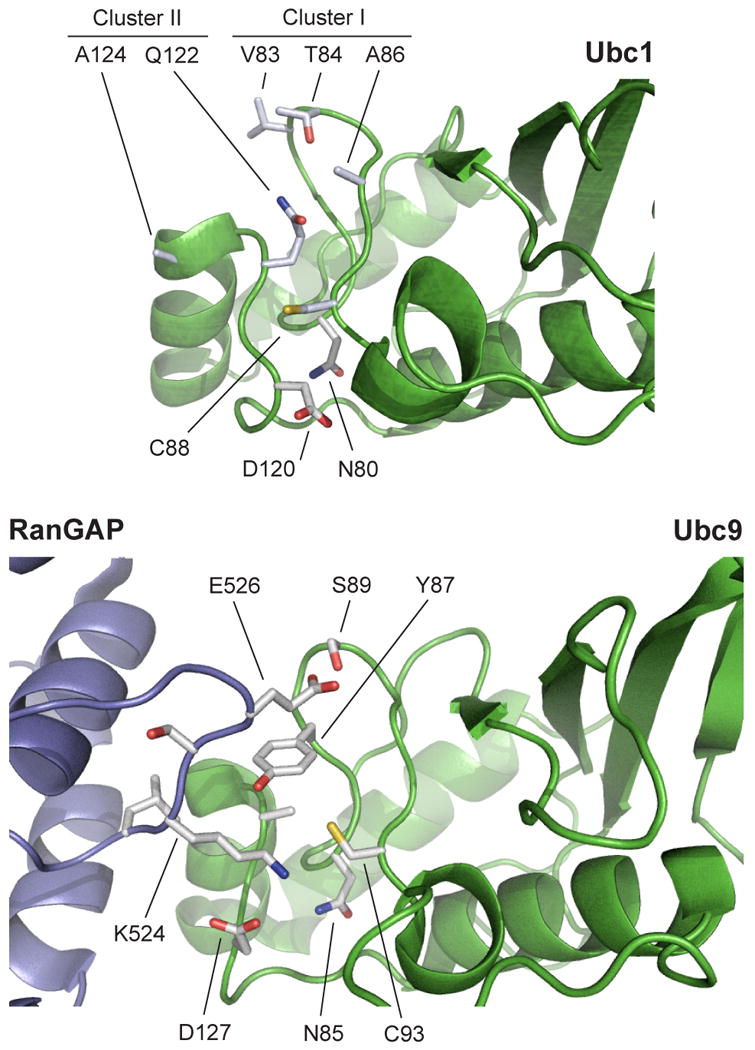
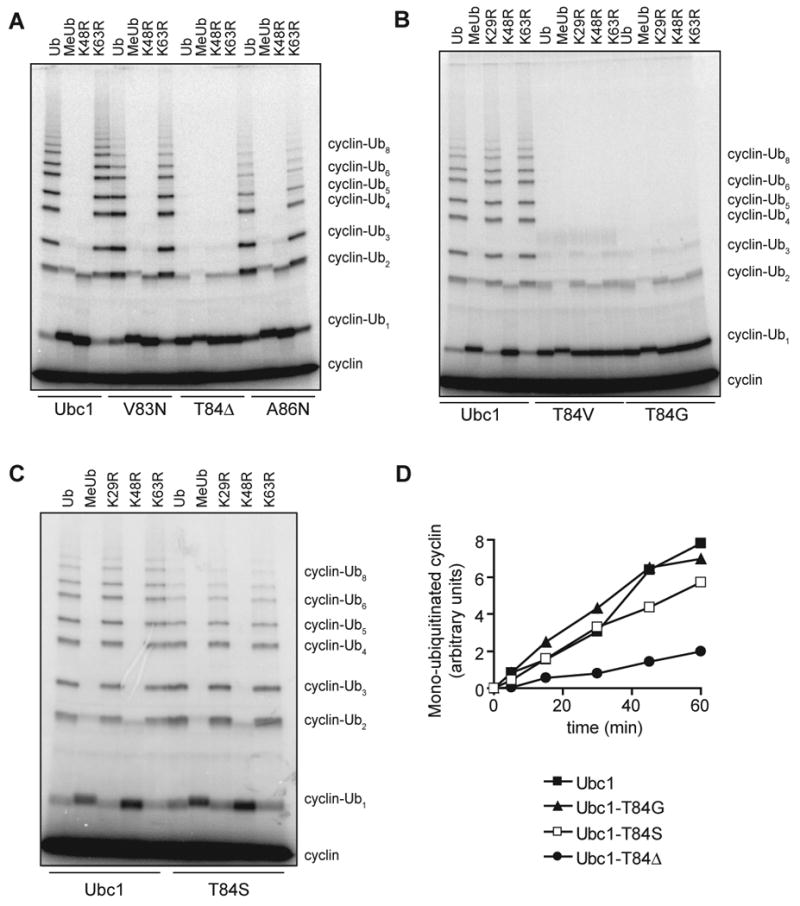
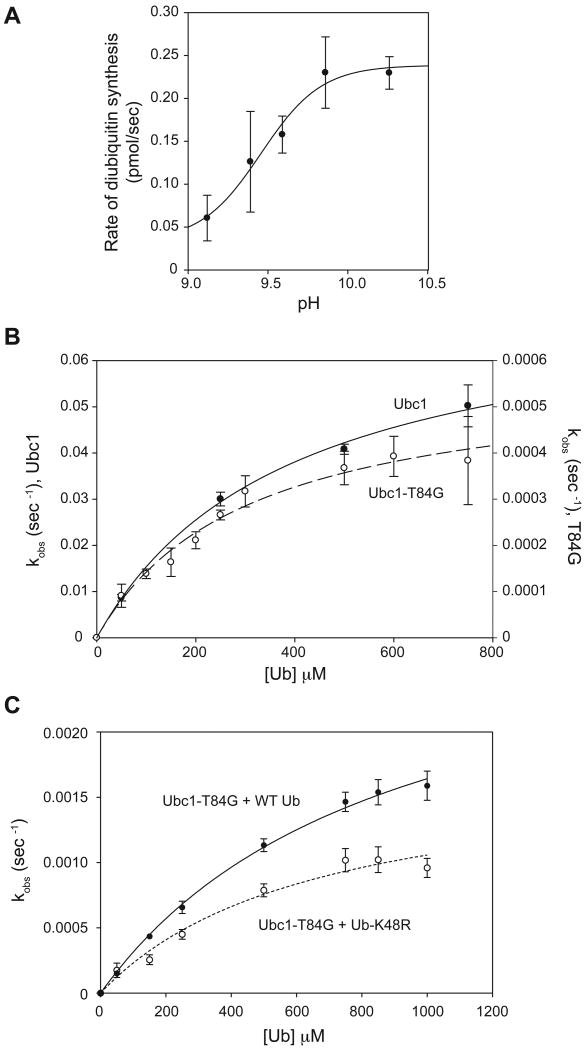
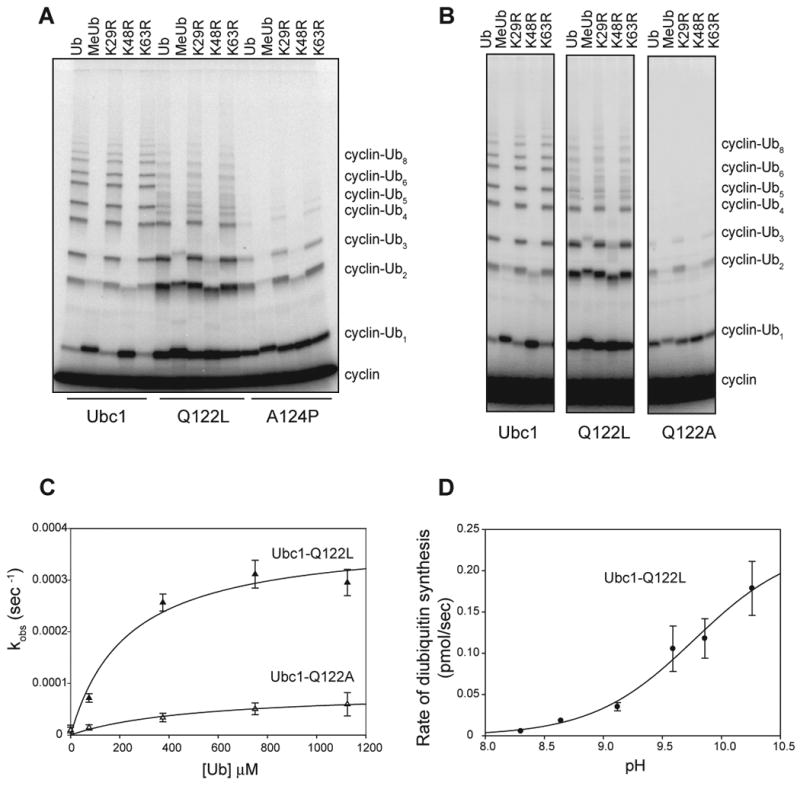
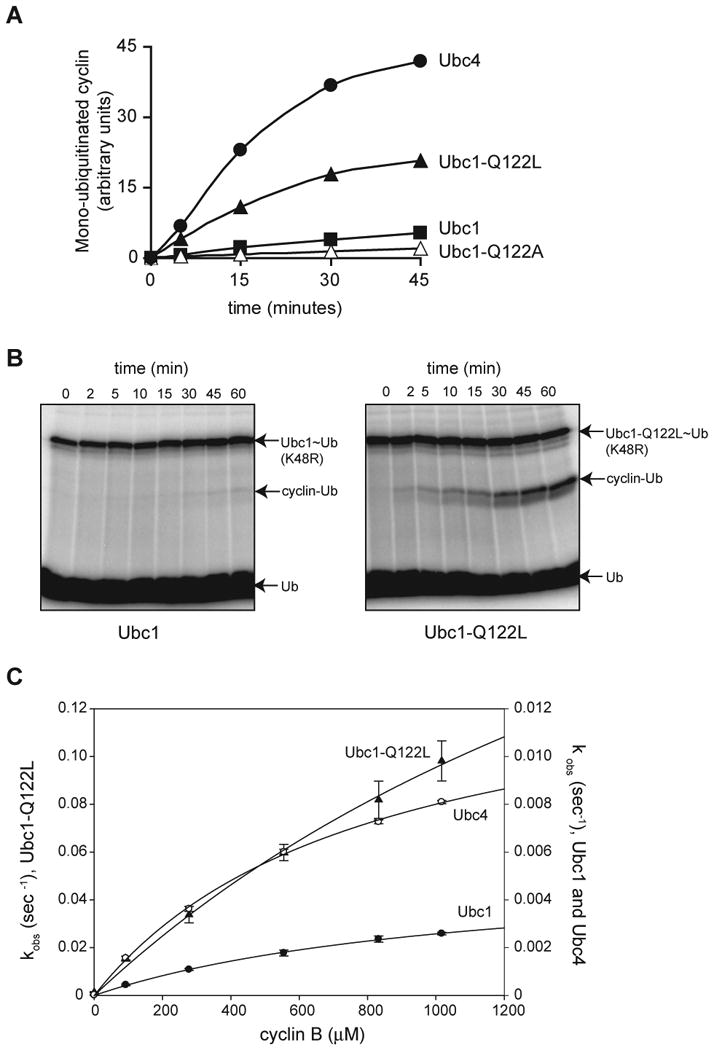
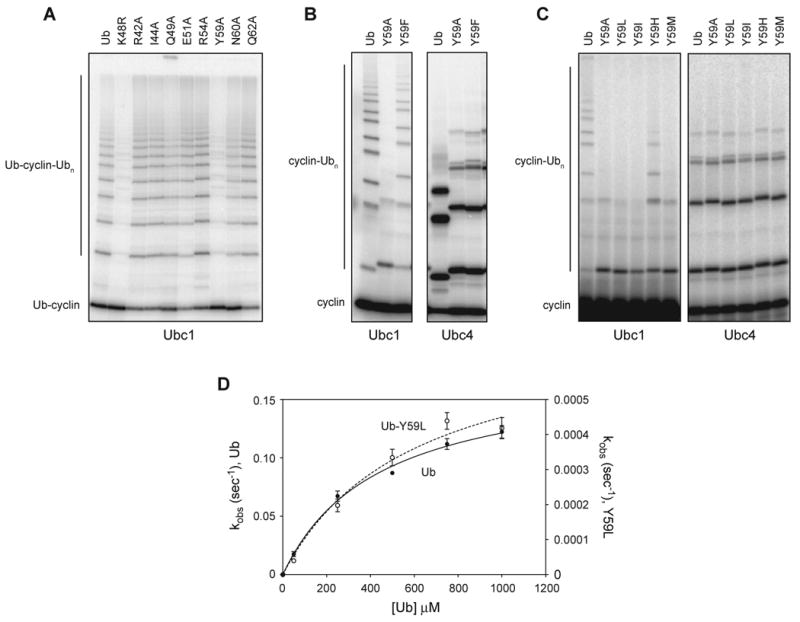
Protein ubiquitination is catalyzed by ubiquitin-conjugating enzymes (E2s) in collaboration with ubiquitin-protein ligases (E3s). This process depends on nucleophilic attack by a substrate lysine on a thioester bond linking the C terminus of ubiquitin to a cysteine in the E2 active site. Different E2 family members display specificity for lysines in distinct contexts. We addressed the mechanistic basis for this lysine selectivity in Ubc1, an E2 that catalyzes the ubiquitination of lysine 48 (K48) in ubiquitin, leading to the formation of K48-linked polyubiquitin chains. We identified a cluster of polar residues near the Ubc1 active site, as well as a residue in ubiquitin itself, that are required for catalysis of K48-specific ubiquitin ligation, but not for general activity toward other lysines. Our results suggest that the active site of Ubc1, as well as the surface of ubiquitin, contains specificity determinants that channel specific lysines to the central residues involved directly in catalysis.
Copyright (c) 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Molecular and structural insight into lysine selection on substrate and ubiquitin lysine 48 by the ubiquitin-conjugating enzyme Cdc34.Cell Cycle. 2013 Jun 1;12(11):1732-44. doi: 10.4161/cc.24818. Epub 2013 May 8. Cell Cycle. 2013. PMID: 23656784 Free PMC article.
-
The UBA domain of conjugating enzyme Ubc1/Ube2K facilitates assembly of K48/K63-branched ubiquitin chains.EMBO J. 2021 Mar 15;40(6):e106094. doi: 10.15252/embj.2020106094. Epub 2021 Feb 12. EMBO J. 2021. PMID: 33576509 Free PMC article.
-
Sequential E2s drive polyubiquitin chain assembly on APC targets.Cell. 2007 Jul 13;130(1):127-39. doi: 10.1016/j.cell.2007.05.027. Cell. 2007. PMID: 17632060
-
The family of ubiquitin-conjugating enzymes (E2s): deciding between life and death of proteins.FASEB J. 2010 Apr;24(4):981-93. doi: 10.1096/fj.09-136259. Epub 2009 Nov 25. FASEB J. 2010. PMID: 19940261 Review.
-
Structural basis of generic versus specific E2-RING E3 interactions in protein ubiquitination.Protein Sci. 2019 Oct;28(10):1758-1770. doi: 10.1002/pro.3690. Epub 2019 Aug 23. Protein Sci. 2019. PMID: 31340062 Free PMC article. Review.
Cited by
-
Analysis of ubiquitin recognition by the HECT ligase E6AP provides insight into its linkage specificity.J Biol Chem. 2019 Apr 12;294(15):6113-6129. doi: 10.1074/jbc.RA118.007014. Epub 2019 Feb 8. J Biol Chem. 2019. PMID: 30737286 Free PMC article.
-
Ubiquitin makes its mark on immune regulation.Immunity. 2010 Dec 14;33(6):843-52. doi: 10.1016/j.immuni.2010.12.007. Immunity. 2010. PMID: 21168777 Free PMC article. Review.
-
Unanchored Ubiquitin Chains, Revisited.Front Cell Dev Biol. 2020 Oct 26;8:582361. doi: 10.3389/fcell.2020.582361. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33195227 Free PMC article. Review.
-
Current perspectives of ubiquitination and SUMOylation in abiotic stress tolerance in plants.Front Plant Sci. 2022 Sep 20;13:993194. doi: 10.3389/fpls.2022.993194. eCollection 2022. Front Plant Sci. 2022. PMID: 36212351 Free PMC article. Review.
-
A comparative study of the cryo-EM structures of Saccharomyces cerevisiae and human anaphase-promoting complex/cyclosome (APC/C).Elife. 2024 Oct 14;13:RP100821. doi: 10.7554/eLife.100821. Elife. 2024. PMID: 39401078 Free PMC article.
References
-
- Bernier-Villamor V, Sampson DA, Matunis MJ, Lima CD. Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell. 2002;108:345–356. - PubMed
-
- Carroll CW, Morgan DO. Enzymology of the Anaphase-Promoting Complex. Meth Enzymol. 2005;398:219–230. - PubMed
-
- Cook WJ, Jeffrey LC, Xu Y, Chau V. Tertiary structures of class I ubiquitin-conjugating enzymes are highly conserved: crystal structure of yeast Ubc4. Biochemistry. 1993;32:13809–13817. - PubMed
-
- DeLano WL. The PyMOL Molecular Graphics System. Palo Alto, CA, USA: DeLano Scientific LLC; 2008. http://www.pymol.org.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
