

Review
doi: 10.1016/j.cell.2010.08.001.
Cellular strategies for regulating DNA supercoiling: a single-molecule perspective
Affiliations
- PMID: 20723754
- PMCID: PMC2997354
- DOI: 10.1016/j.cell.2010.08.001
Item in Clipboard
Review
Cellular strategies for regulating DNA supercoiling: a single-molecule perspective
Cell.
.
Abstract
Entangling and twisting of cellular DNA (i.e., supercoiling) are problems inherent to the helical structure of double-stranded DNA. Supercoiling affects transcription, DNA replication, and chromosomal segregation. Consequently the cell must fine-tune supercoiling to optimize these key processes. Here, we summarize how supercoiling is generated and review experimental and theoretical insights into supercoil relaxation. We distinguish between the passive dissipation of supercoils by diffusion and the active removal of supercoils by topoisomerase enzymes. We also review single-molecule studies that elucidate the timescales and mechanisms of supercoil removal.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
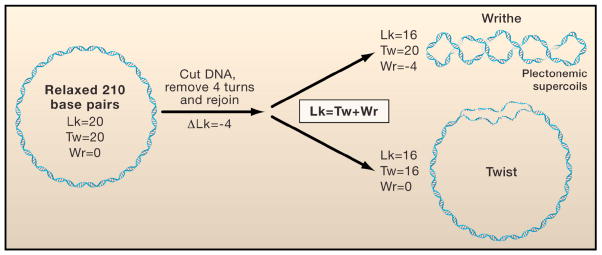
The topology of a double-stranded DNA is described by its linking number (Lk), which is the sum of twist (Tw) and writhe (Wr). (Left) A torsionally relaxed DNA molecule with a length of 210 basepairs contains 20 turns (10.5 bp/turn) or Tw=20. Hypothetically, if the DNA were cut, then one end was twisted by four turns in the direction opposite to the natural helicity of the DNA, and subsequently resealed, the resulting linking number of the DNA would equal Lk=20−4=16. (Right) The upper and lower panels show the topology of the DNA molecule when the removal of these turns is at the expense of twist (Tw=20−4=16) and writhe (Wr=0−4=−4), respectively.
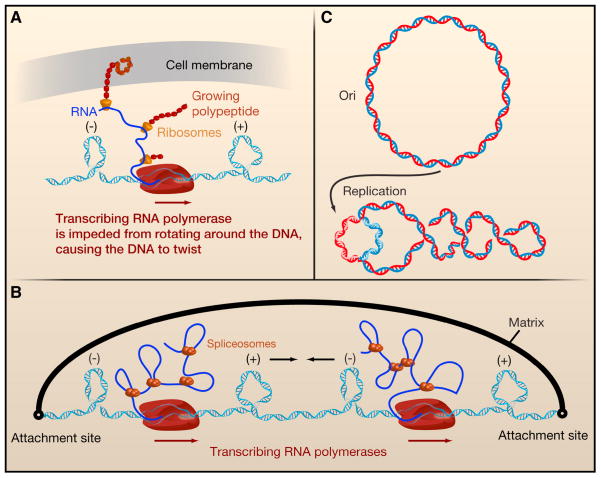
A. When RNA polymerase (RNAP) is prevented from rotating along the helical axis of the DNA during transcription, positive and negative supercoils accumulate ahead and behind the enzyme, respectively. Multiple factors impede the rotation of RNAP by increasing its hydrodynamic drag. In bacteria, these factors include the nascent RNA strand (blue solid line), ribosomes on the mRNA (yellow), and even the growing peptide itself. Furthermore, the nascent protein might insert itself into the cell membrane (gray sheet), providing an anchor point. B. In eukaryotes, the nascent RNA and its processing factors, such as spliceosomes, increase the rotational drag on RNAP and impede its rotation around DNA’s helical axis, leading to supercoiling behind and ahead of the enzyme. When tandem genes are transcribed, RNAP complexes progress in the same direction on duplex DNA. The DNA domain between them contains both negative and positive supercoils that could diffuse towards each other and subsequently annihilate. The rate at which this process occurs depends on the timescales at which DNA can spin around its own helical axis and the axis defined by plectonemes. C. When a circular DNA (top) is replicated, two origins move in opposite directions, unwinding the parental DNA. By conservation of linking number, this generates positive supercoils ahead of the forks (bottom).
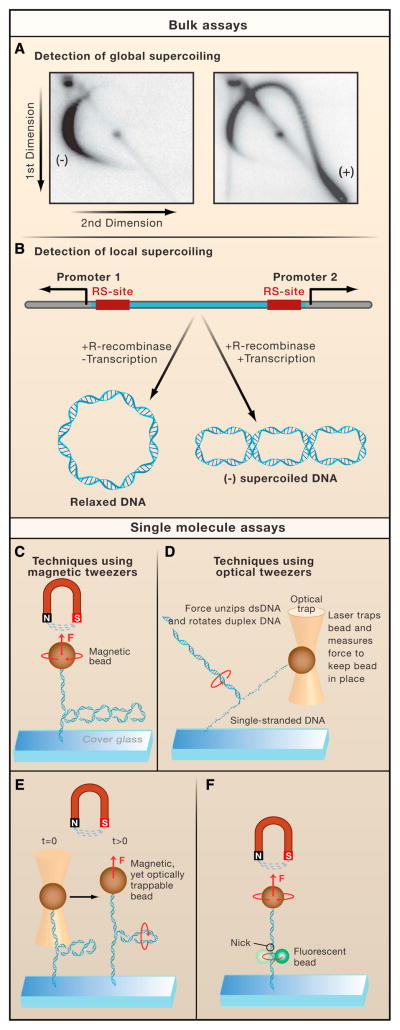
A. Global supercoiling can be detected using two-dimensional gel electrophoresis, in which plasmid DNA is first resolved in the gel in one direction, followed by electrophoresis in a perpendicular direction in the presence of an increased concentration of an intercalating agent. As a result, an arc-shaped pattern emerges, consisting of spots where the degree of supercoiling (i.e., linking number or Lk) increases in a clockwise orientation (Left panel). Plasmid DNA isolated from yeast cells expressing TopIB yields a population of negatively supercoiled topoisomers because the DNA is wrapped around nucleosomes in vivo (Right panel). By contrast, in cells that lack TopIB or TopII but express E. coli TopIA, the preferential relaxation of negatively supercoiled DNA induced by transcription, results in the accumulation of positively supercoiled DNA (right panel). B. The ring excision assay can detect local supercoiling in a predefined region on the DNA. When a recombinase excises a stretch of DNA between two recombinase sites (RS), the resulting ring of DNA retains the supercoil density it had when it was still integrated. This assay was used to characterize the negative supercoils generated between two RNAP complexes moving away from divergent promoters. C. Magnetic tweezers exert a magnetic force (F) on a magnetic bead, which is attached to a double-stranded DNA molecule. The other end of the molecule is linked to a microscope coverslip. By translating the magnets up or down, one increases or decreases the pulling force, respectively. The DNA can be supercoiled by twisting the magnets. D. A focused laser can trap a transparent bead at a fixed position in space and measure the force exerted on the bead. These optical tweezers can measure the rotational drag of non-plectonemic duplex DNA with one strand tethered to the glass slide and the other strand held stationary in the optical trap. As the stage is moved away from the trap, the duplex DNA starts to unzip, forcing the double-stranded portion of the DNA to spin around its axis. The optical trap measures the force required to start unzipping the DNA. E. A combination of magnetic and optical tweezers has been used to measure the rotational drag of plectonemic DNA. Here, the pulling force on a supercoiled DNA molecule is suddenly increased. During this process, the plectonemes will rotate as indicated, generating viscous drag that opposes the motion of the bead. F. The rotor bead assay consists of a standard magnetic tweezers setup augmented with a strategically positioned small fluorescent “rotor bead”. This rotor bead serves as an indicator for the rotational motion of DNA as it spins along the axis of the DNA concurrently with the DNA.
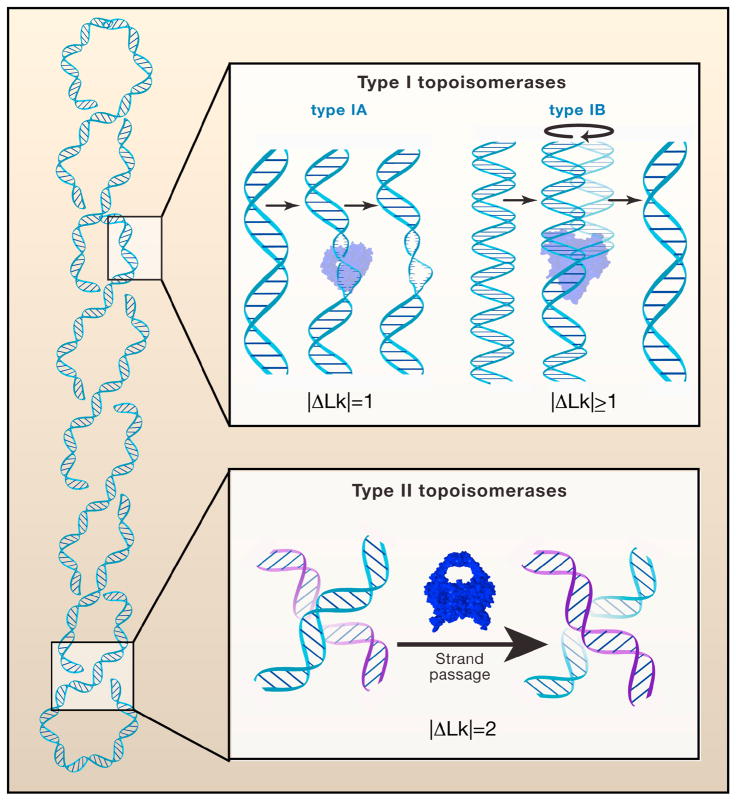
(Top) Type I topoisomerases cleave a single-strand of DNA and relax a supercoil by either passing the other strand through an enzyme-DNA linked intermediate (type IA enzymes) or by a strand-swivel mechanism (type IB enzymes). (Bottom) Type II topoisomerases cleave duplex DNA and then relax the supercoil by passing a second duplex DNA through the transient enzyme-DNA linked intermediate.
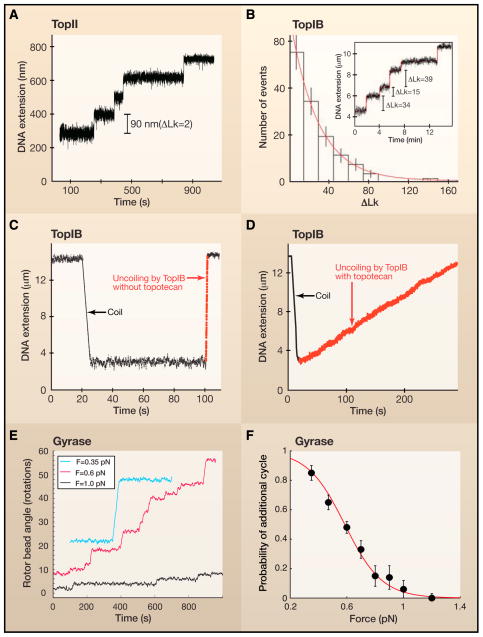
A. In a magnetic tweezer experiment, the length or extension of the DNA provides a measure of its supercoil density at a fixed force. In the presence of TopII, the DNA extension increases with time in equally-sized steps of 90 nm which is equivalent to the removal of two supercoils for each catalytic cycle. B. In contrast to TopII, TopIB does not remove a fixed number of supercoils with each catalytic event (inset). Rather, the distribution of the number of supercoils removed is exponential, with a mean far above unity. This result agrees with the proposed swivel mechanism for supercoil relaxation by TopIB. C. The instantaneous relaxation rate of supercoils by TopIB in the absence of the chemotherapeutic drug topotecan is relatively fast. In this trace, the rate is ~100 supercoils per second (dashed red line) D. The chemotherapeutic drug topotecan slows down relaxation dramatically, to the extent that the typical rates are now ~5 supercoils per second. E. Rotor bead assays showed that DNA gyrase introduces two supercoils with each catalytic cycle. F. The processivity of gyrase activity decreases markedly with increasing force, suggesting the presence of a force-sensitive step in the enzyme’s catalytic cycle. It is thought that the enzyme performs work against the externally applied force when wrapping a DNA segment around itself. The solid line is a fit to a model that includes competition between this wrapping step and enzyme dissociation from the DNA.
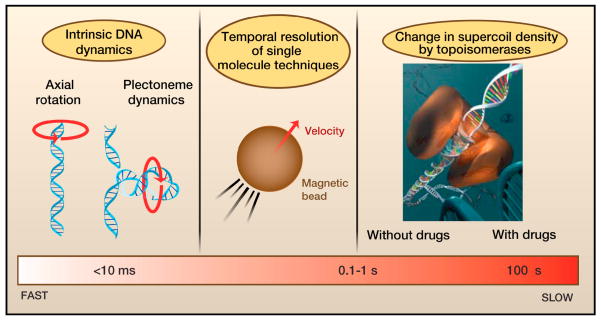
From fast (left) to slow (right): The rotation of DNA around its own axis (as described by the “speedometer” model) and the rotation of plectonemes about their central axis are estimated to occur on timescales faster than 100 μs per one unit change in Lk (i.e., |ΔLk|=1). The temporal resolution for the measurement of supercoil removal using single-molecule techniques is ~10 ms and is limited by the dynamics of the force sensors (e.g. micron-sized beads). Much slower processes are the addition or removal of a few supercoils by enzymes, such as gyrase and Type IB topoisomerases (typically ~10 ms per supercoil). For the latter, inhibitor drugs have been shown to dramatically slow down supercoil removal to approximately 0.2 s per supercoil for the removal of positive supercoils.
Similar articles
-
Control of bacterial DNA supercoiling.Mol Microbiol. 1992 Feb;6(4):425-33. doi: 10.1111/j.1365-2958.1992.tb01486.x. Mol Microbiol. 1992. PMID: 1313943 Review.
-
[DNA supercoiling and topoisomerases in Escherichia coli].Rev Latinoam Microbiol. 1995 Jul-Sep;37(3):291-304. Rev Latinoam Microbiol. 1995. PMID: 8850348 Review. Spanish.
-
A multiscale dynamic model of DNA supercoil relaxation by topoisomerase IB.Biophys J. 2011 Apr 20;100(8):2016-23. doi: 10.1016/j.bpj.2011.03.003. Biophys J. 2011. PMID: 21504738 Free PMC article.
-
Rotation of DNA around intact strand in human topoisomerase I implies distinct mechanisms for positive and negative supercoil relaxation.Nucleic Acids Res. 2005 Nov 27;33(20):6621-34. doi: 10.1093/nar/gki935. Print 2005. Nucleic Acids Res. 2005. PMID: 16314322 Free PMC article.
-
DNA supercoiling during ATP-dependent DNA translocation by the type I restriction enzyme EcoAI.J Mol Biol. 2000 Jan 28;295(4):1089-99. doi: 10.1006/jmbi.1999.3414. J Mol Biol. 2000. PMID: 10656812
Cited by
-
DNA Manipulation and Single-Molecule Imaging.Molecules. 2021 Feb 17;26(4):1050. doi: 10.3390/molecules26041050. Molecules. 2021. PMID: 33671359 Free PMC article. Review.
-
High-Resolution Genome-Wide Maps Reveal Widespread Presence of Torsional Insulation.bioRxiv [Preprint]. 2025 Jan 4:2024.10.11.617876. doi: 10.1101/2024.10.11.617876. bioRxiv. 2025. PMID: 39416127 Free PMC article. Preprint.
-
Psoralen mapping reveals a bacterial genome supercoiling landscape dominated by transcription.Nucleic Acids Res. 2022 May 6;50(8):4436-4449. doi: 10.1093/nar/gkac244. Nucleic Acids Res. 2022. PMID: 35420137 Free PMC article.
-
Application of Plasmid Engineering to Enhance Yield and Quality of Plasmid for Vaccine and Gene Therapy.Bioengineering (Basel). 2019 Jun 19;6(2):54. doi: 10.3390/bioengineering6020054. Bioengineering (Basel). 2019. PMID: 31248216 Free PMC article.
-
The dynamic interplay between DNA topoisomerases and DNA topology.Biophys Rev. 2016 Nov;8(Suppl 1):101-111. doi: 10.1007/s12551-016-0240-8. Epub 2016 Nov 14. Biophys Rev. 2016. PMID: 28510219 Free PMC article. Review.
References
-
- Bishop AI, Nieminen TA, Heckenberg NR, Rubinsztein-Dunlop H. Optical application and measurement of torque on microparticles of isotropic nonabsorbing material. Phys Rev A. 2003;68:033802.
-
- Bohbot-Raviv Y, Zhao WZ, Feingold M, Wiggins CH, Granek R. Relaxation dynamics of semiflexible polymers. Phys Rev Lett. 2004;92:098101. - PubMed
-
- Brown PO, Cozzarelli NR. Sign Inversion Mechanism for Enzymatic Supercoiling of DNA. Science. 1979;206:1081–1083. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
