

Dynamic chromatin organization during foregut development mediated by the organ selector gene PHA-4/FoxA
- PMID: 20714352
- PMCID: PMC2920861
- DOI: 10.1371/journal.pgen.1001060
Dynamic chromatin organization during foregut development mediated by the organ selector gene PHA-4/FoxA
Abstract
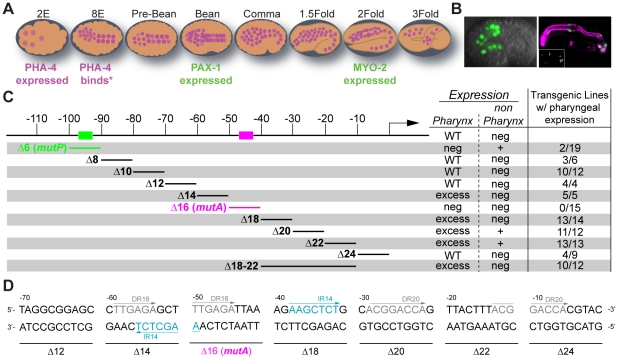
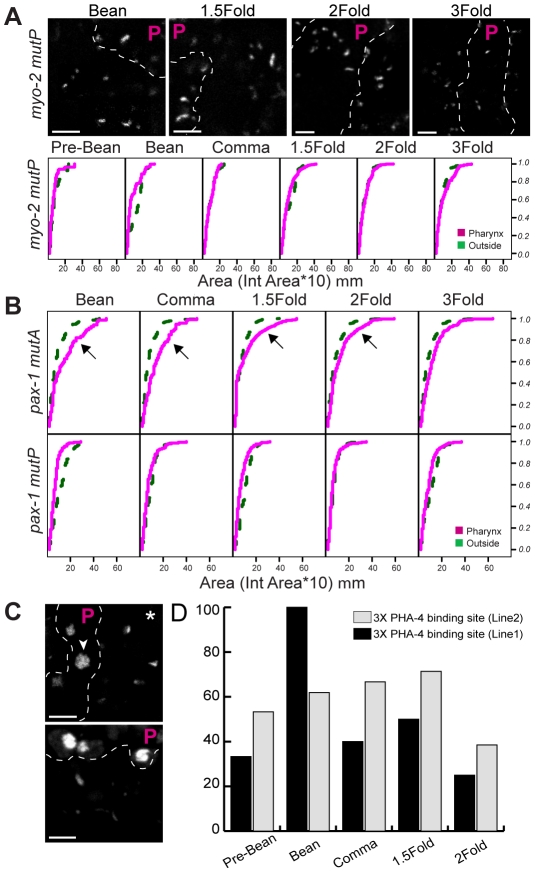
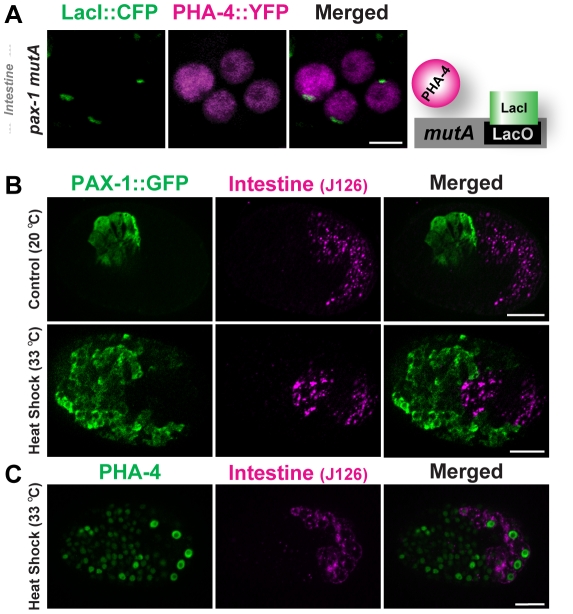
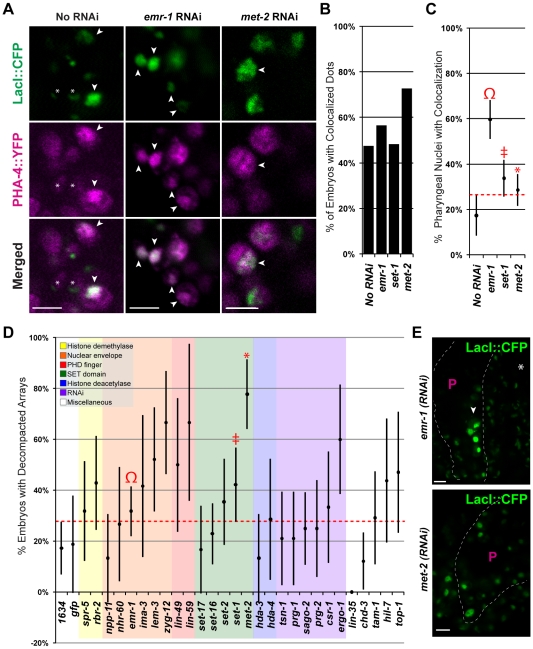
Central regulators of cell fate, or selector genes, establish the identity of cells by direct regulation of large cohorts of genes. In Caenorhabditis elegans, foregut (or pharynx) identity relies on the FoxA transcription factor PHA-4, which activates different sets of target genes at various times and in diverse cellular environments. An outstanding question is how PHA-4 distinguishes between target genes for appropriate transcriptional control. We have used the Nuclear Spot Assay and GFP reporters to examine PHA-4 interactions with target promoters in living embryos and with single cell resolution. While PHA-4 was found throughout the digestive tract, binding and activation of pharyngeally expressed promoters was restricted to a subset of pharyngeal cells and excluded from the intestine. An RNAi screen of candidate nuclear factors identified emerin (emr-1) as a negative regulator of PHA-4 binding within the pharynx, but emr-1 did not modulate PHA-4 binding in the intestine. Upon promoter association, PHA-4 induced large-scale chromatin de-compaction, which, we hypothesize, may facilitate promoter access and productive transcription. Our results reveal two tiers of PHA-4 regulation. PHA-4 binding is prohibited in intestinal cells, preventing target gene expression in that organ. PHA-4 binding within the pharynx is limited by the nuclear lamina component EMR-1/emerin. The data suggest that association of PHA-4 with its targets is a regulated step that contributes to promoter selectivity during organ formation. We speculate that global re-organization of chromatin architecture upon PHA-4 binding promotes competence of pharyngeal gene transcription and, by extension, foregut development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
pha-4 is Ce-fkh-1, a fork head/HNF-3alpha,beta,gamma homolog that functions in organogenesis of the C. elegans pharynx.Development. 1998 Jun;125(12):2171-80. doi: 10.1242/dev.125.12.2171. Development. 1998. PMID: 9584117
-
Temporal regulation of foregut development by HTZ-1/H2A.Z and PHA-4/FoxA.PLoS Genet. 2006 Sep 29;2(9):e161. doi: 10.1371/journal.pgen.0020161. Epub 2006 Aug 11. PLoS Genet. 2006. PMID: 17009877 Free PMC article.
-
PHA-4/FoxA cooperates with TAM-1/TRIM to regulate cell fate restriction in the C. elegans foregut.Dev Biol. 2007 Mar 15;303(2):611-24. doi: 10.1016/j.ydbio.2006.11.042. Epub 2006 Dec 2. Dev Biol. 2007. PMID: 17250823 Free PMC article.
-
The molecular basis of organ formation: insights from the C. elegans foregut.Annu Rev Cell Dev Biol. 2009;25:597-628. doi: 10.1146/annurev.cellbio.24.110707.175411. Annu Rev Cell Dev Biol. 2009. PMID: 19575642 Free PMC article. Review.
-
Development of Caenorhabditis elegans pharynx, with emphasis on its nervous system.Acta Pharmacol Sin. 2005 Apr;26(4):396-404. doi: 10.1111/j.1745-7254.2005.00070.x. Acta Pharmacol Sin. 2005. PMID: 15780187 Review.
Cited by
-
A high-content imaging approach to profile C. elegans embryonic development.Development. 2019 Apr 11;146(7):dev174029. doi: 10.1242/dev.174029. Development. 2019. PMID: 30890570 Free PMC article.
-
Pioneer transcription factors shape the epigenetic landscape.J Biol Chem. 2018 Sep 7;293(36):13795-13804. doi: 10.1074/jbc.R117.001232. Epub 2018 Mar 5. J Biol Chem. 2018. PMID: 29507097 Free PMC article. Review.
-
Pioneer transcription factors, chromatin dynamics, and cell fate control.Curr Opin Genet Dev. 2016 Apr;37:76-81. doi: 10.1016/j.gde.2015.12.003. Epub 2016 Jan 27. Curr Opin Genet Dev. 2016. PMID: 26826681 Free PMC article. Review.
-
Ce-emerin and LEM-2: essential roles in Caenorhabditis elegans development, muscle function, and mitosis.Mol Biol Cell. 2012 Feb;23(4):543-52. doi: 10.1091/mbc.E11-06-0505. Epub 2011 Dec 14. Mol Biol Cell. 2012. PMID: 22171324 Free PMC article.
-
Transcriptional coactivator PGC-1α contributes to decidualization by forming a histone-modifying complex with C/EBPβ and p300.J Biol Chem. 2022 May;298(5):101874. doi: 10.1016/j.jbc.2022.101874. Epub 2022 Mar 28. J Biol Chem. 2022. PMID: 35358514 Free PMC article.
References
-
- Mann RS, Carroll SB. Molecular mechanisms of selector gene function and evolution. Curr Opin Genet Dev. 2002;12:592–600. - PubMed
-
- Gaudet J, Mango SE. Regulation of organogenesis by the Caenorhabditis elegans FoxA protein PHA-4. Science. 2002;295:821–825. - PubMed
-
- Furlong EE, Andersen EC, Null B, White KP, Scott MP. Patterns of gene expression during Drosophila mesoderm development. Science. 2001;293:1629–1633. - PubMed
-
- Tapscott SJ. The circuitry of a master switch: Myod and the regulation of skeletal muscle gene transcription. Development. 2005;132:2685–2695. - PubMed
-
- Liang Z, Biggin MD. Eve and ftz regulate a wide array of genes in blastoderm embryos: the selector homeoproteins directly or indirectly regulate most genes in Drosophila. Development. 1998;125:4471–4482. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
