

Review
doi: 10.1083/jcb.201001138.
Review series: The cell biology of hearing
Affiliations
- PMID: 20624897
- PMCID: PMC2911669
- DOI: 10.1083/jcb.201001138
Item in Clipboard
Review
Review series: The cell biology of hearing
J Cell Biol.
.
Abstract
Mammals have an astonishing ability to sense and discriminate sounds of different frequencies and intensities. Fundamental for this process are mechanosensory hair cells in the inner ear that convert sound-induced vibrations into electrical signals. The study of genes that are linked to deafness has provided insights into the cell biological mechanisms that control hair cell development and their function as mechanosensors.
Figures
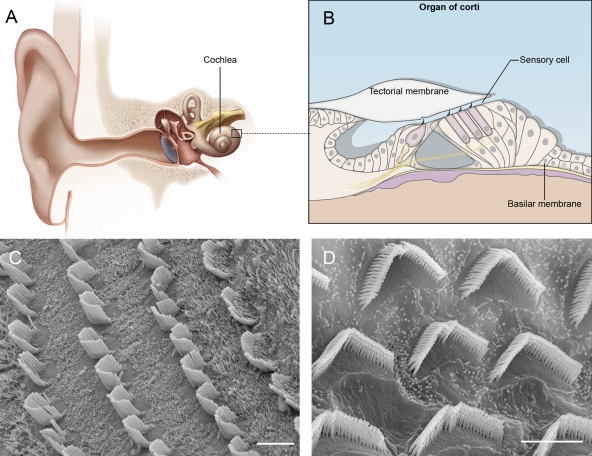
The auditory sense organ. (A) Diagram of the auditory sense organ highlighting the snail-shaped cochlea. (B) Diagram of the organ of Corti. (C) Scanning electron micrographs of hair bundles in the cochlea after removal of the tectorial membrane. Three rows of OHCs are shown at the left, one row of IHCs at the right. (D) Higher magnification view of OHCs. Bars, 5 µm.
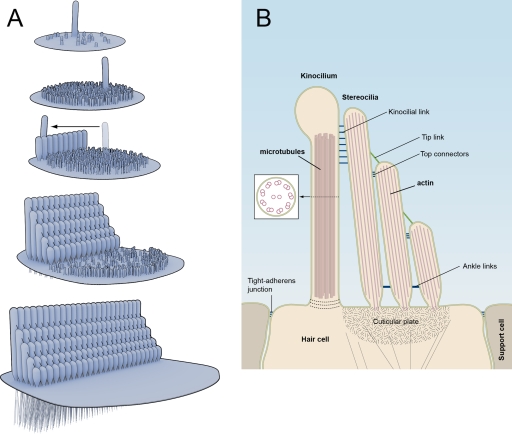
Hair bundle development and structure. (A) Diagram of sequential stages of hair bundle development. At the onset, the apical hair cell surface contains microvilli and one kinocilium. The microvilli grow in length. The kinocilium moves to the lateral edge of the hair cell. Some microvilli elongate to form stereocilia of graded heights. (B) Cross section through a hair bundle and apical hair cell surface indicating the kinocilium, stereocilia, and cuticular plate. Some of the linkages in hair bundles are highlighted.
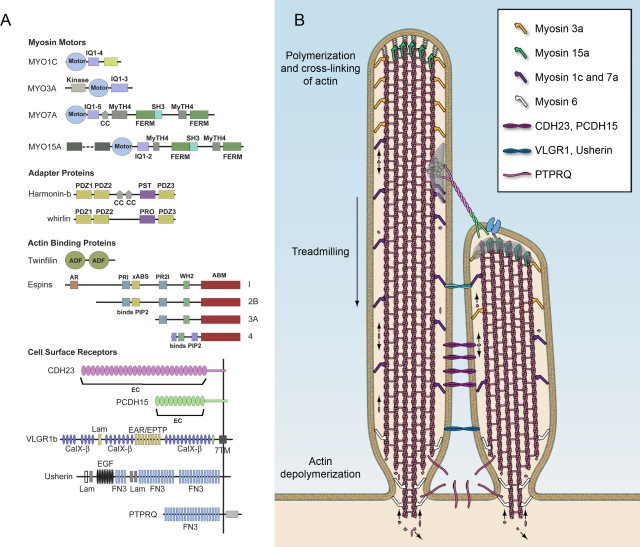
Hair bundle proteins. (A) Domain structure of proteins discussed in the text. Abbreviations: CC, coiled-coil domain; FERM, protein 4.1, ezrin, radixin, moesin domain; IQ, calmodulin-binding IQ domain; MyTH4, myosin tail homology 4 domain; PDZ, PSD95/SAP90, Discs large, zonula occludens-1 domain; PST, proline, serine, threonine-rich domain; PRO, proline-rich domain; SH3, src homology 3 domain; ADF, actin-depolymerization factor; AR, ankyrin-like repeat; PR, proline-rich peptide; ABS, F-actin–binding site; WH, WASP homology 2 domain; ABM, actin bundling module; EC, extracellular cadherin repeat; CalX-β, Ca2+-binding calcium enhancer β modules; Lam, Laminin GL or NT domain; EAR/EPTP, putative β-propeller folding domain; EGF, laminin-type epidermal growth factor–like domain; FN3, fibronectin type 3 repeat. (B) Diagram of two stereocilia indicating the distribution of some of the molecules discussed in the text.
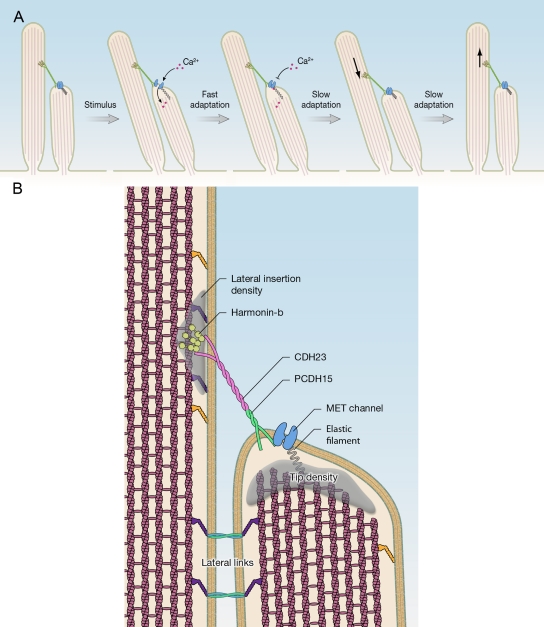
Hair bundles and mechanotransduction. Model of transduction and adaptation. Deflection of hair bundles in the direction of the longest stereocilia leads to the opening of transduction channels at the lower ends of tip links. Ca2+ enters the transduction channel and binds to the channel or a side near the channel and leads to channel closure (fast adaptation). The adaptation motor at the upper end of tip links subsequently detaches from the actin cytoskeleton and slides down the stereocilium, leading to release of tension in the transduction machinery (slipping phase of slow adaptation). Next, the motor complex climbs up the stereocilium, reestablishing tension (climbing phase of slow adaptation). (B) Molecular components of the mechanotransduction complex in stereocilia.
Similar articles
-
Tmc proteins are essential for zebrafish hearing where Tmc1 is not obligatory.Hum Mol Genet. 2020 Jul 29;29(12):2004-2021. doi: 10.1093/hmg/ddaa045. Hum Mol Genet. 2020. PMID: 32167554 Free PMC article.
-
[Molecular biology of hearing].Laryngorhinootologie. 2011 Mar;90 Suppl 1:S22-34. doi: 10.1055/s-0030-1270444. Epub 2011 Apr 26. Laryngorhinootologie. 2011. PMID: 21523631 Review. German.
-
Hair cells, plasma membrane Ca²⁺ ATPase and deafness.Int J Biochem Cell Biol. 2012 May;44(5):679-83. doi: 10.1016/j.biocel.2012.02.006. Epub 2012 Feb 13. Int J Biochem Cell Biol. 2012. PMID: 22349217 Review.
-
Kiaa1024L/Minar2 is essential for hearing by regulating cholesterol distribution in hair bundles.Elife. 2022 Nov 1;11:e80865. doi: 10.7554/eLife.80865. Elife. 2022. PMID: 36317962 Free PMC article.
-
Actin-bundling protein TRIOBP forms resilient rootlets of hair cell stereocilia essential for hearing.Cell. 2010 May 28;141(5):786-98. doi: 10.1016/j.cell.2010.03.049. Cell. 2010. PMID: 20510926 Free PMC article.
Cited by
-
Developmental and architectural principles of the lateral-line neural map.Front Neural Circuits. 2013 Mar 26;7:47. doi: 10.3389/fncir.2013.00047. eCollection 2013. Front Neural Circuits. 2013. PMID: 23532704 Free PMC article. Review.
-
The Cell Adhesion Molecules Roughest, Hibris, Kin of Irre and Sticks and Stones Are Required for Long Range Spacing of the Drosophila Wing Disc Sensory Sensilla.PLoS One. 2015 Jun 8;10(6):e0128490. doi: 10.1371/journal.pone.0128490. eCollection 2015. PLoS One. 2015. PMID: 26053791 Free PMC article.
-
Constitutive activation of DIA1 (DIAPH1) via C-terminal truncation causes human sensorineural hearing loss.EMBO Mol Med. 2016 Nov 2;8(11):1310-1324. doi: 10.15252/emmm.201606609. Print 2016 Nov. EMBO Mol Med. 2016. PMID: 27707755 Free PMC article.
-
Molecular biology of hearing.GMS Curr Top Otorhinolaryngol Head Neck Surg. 2011;10:Doc06. doi: 10.3205/cto000079. Epub 2012 Apr 26. GMS Curr Top Otorhinolaryngol Head Neck Surg. 2011. PMID: 22558056 Free PMC article.
-
The Emerging Role of Sperm-Associated Antigen 6 Gene in the Microtubule Function of Cells and Cancer.Mol Ther Oncolytics. 2019 Sep 10;15:101-107. doi: 10.1016/j.omto.2019.08.011. eCollection 2019 Dec 20. Mol Ther Oncolytics. 2019. PMID: 31660426 Free PMC article. Review.
References
-
- Adato A., Lefèvre G., Delprat B., Michel V., Michalski N., Chardenoux S., Weil D., El-Amraoui A., Petit C. 2005a. Usherin, the defective protein in Usher syndrome type IIA, is likely to be a component of interstereocilia ankle links in the inner ear sensory cells. Hum. Mol. Genet. 14:3921–3932 10.1093/hmg/ddi416 - DOI - PubMed
-
- Ahmed Z.M., Goodyear R., Riazuddin S., Lagziel A., Legan P.K., Behra M., Burgess S.M., Lilley K.S., Wilcox E.R., Riazuddin S., et al. 2006. The tip-link antigen, a protein associated with the transduction complex of sensory hair cells, is protocadherin-15. J. Neurosci. 26:7022–7034 10.1523/JNEUROSCI.1163-06.2006 - DOI - PMC - PubMed
-
- Alagramam K.N., Murcia C.L., Kwon H.Y., Pawlowski K.S., Wright C.G., Woychik R.P. 2001. The mouse Ames waltzer hearing-loss mutant is caused by mutation of Pcdh15, a novel protocadherin gene. Nat. Genet. 27:99–102 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
