

Myeloid cells, BAFF, and IFN-gamma establish an inflammatory loop that exacerbates autoimmunity in Lyn-deficient mice
- PMID: 20624892
- PMCID: PMC2916124
- DOI: 10.1084/jem.20100086
Myeloid cells, BAFF, and IFN-gamma establish an inflammatory loop that exacerbates autoimmunity in Lyn-deficient mice
Abstract
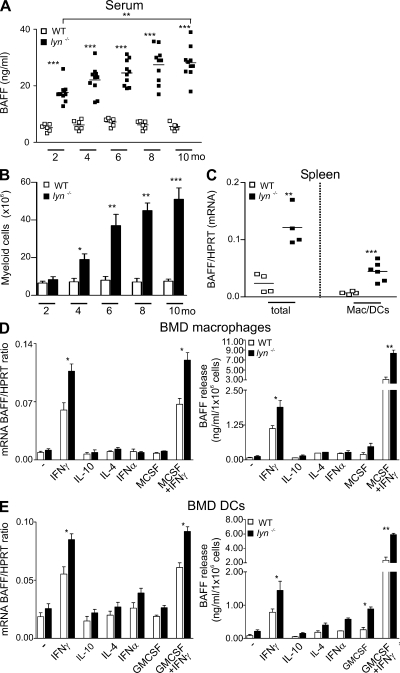
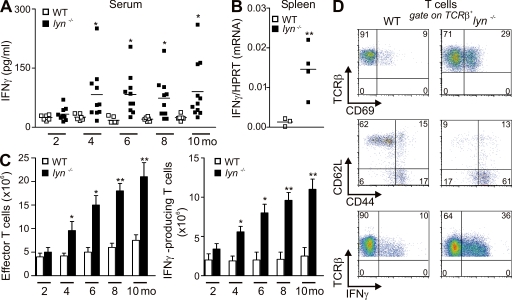
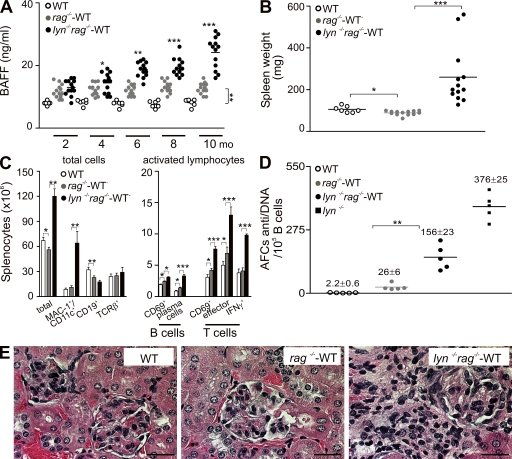
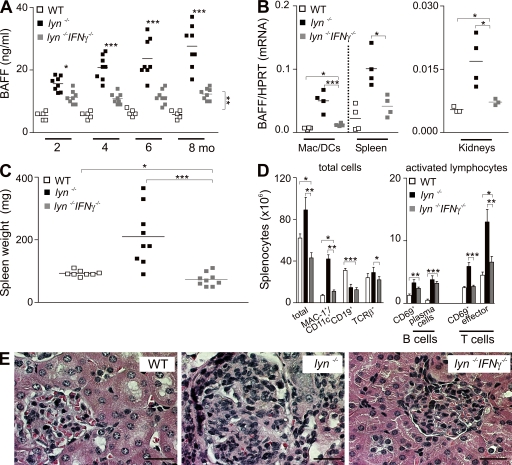
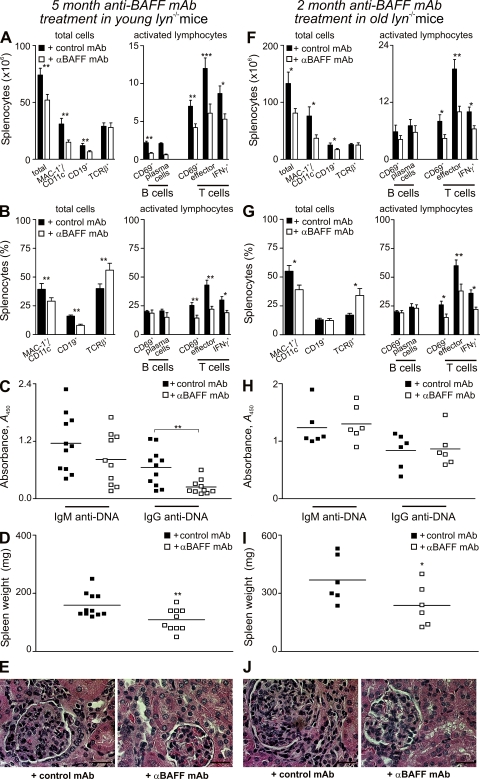
Autoimmunity is traditionally attributed to altered lymphoid cell selection and/or tolerance, whereas the contribution of innate immune cells is less well understood. Autoimmunity is also associated with increased levels of B cell-activating factor of the TNF family (BAFF; also known as B lymphocyte stimulator), a cytokine that promotes survival of self-reactive B cell clones. We describe an important role for myeloid cells in autoimmune disease progression. Using Lyn-deficient mice, we show that overproduction of BAFF by hyperactive myeloid cells contributes to inflammation and autoimmunity in part by acting directly on T cells to induce the release of IFN-gamma. Genetic deletion of IFN-gamma or reduction of BAFF activity, achieved by either reducing myeloid cell hyperproduction or by treating with an anti-BAFF monoclonal antibody, reduced disease development in lyn(-/-) mice. The increased production of IFN-gamma in lyn(-/-) mice feeds back on the myeloid cells to further stimulate BAFF release. Expression of BAFF receptor on T cells was required for their full activation and IFN-gamma release. Overall, our data suggest that the reciprocal production of BAFF and IFN-gamma establishes an inflammatory loop between myeloid cells and T cells that exacerbates autoimmunity in this model. Our findings uncover an important pathological role of BAFF in autoimmune disorders.
Figures



Similar articles
-
BAFF regulates activation of self-reactive T cells through B-cell dependent mechanisms and mediates protection in NOD mice.Eur J Immunol. 2014 Apr;44(4):983-93. doi: 10.1002/eji.201344186. Epub 2014 Feb 16. Eur J Immunol. 2014. PMID: 24435807
-
The receptor tyrosine kinase MerTK regulates dendritic cell production of BAFF.Autoimmunity. 2009 Mar;42(3):183-97. doi: 10.1080/08916930802668586. Autoimmunity. 2009. PMID: 19301199 Free PMC article.
-
Development of autoimmune nephritis in genetically asplenic and splenectomized BAFF transgenic mice.J Autoimmun. 2011 Mar;36(2):125-34. doi: 10.1016/j.jaut.2010.12.002. Epub 2011 Jan 7. J Autoimmun. 2011. PMID: 21216131
-
Regulation of B cell self-tolerance by BAFF.Semin Immunol. 2006 Oct;18(5):276-83. doi: 10.1016/j.smim.2006.04.003. Epub 2006 Aug 17. Semin Immunol. 2006. PMID: 16916609 Review.
-
B cells flying solo.Immunol Cell Biol. 2008 Jan;86(1):40-6. doi: 10.1038/sj.icb.7100142. Immunol Cell Biol. 2008. PMID: 18172443 Review.
Cited by
-
Granulocyte colony-stimulating factor is not pathogenic in lupus nephritis.Immun Inflamm Dis. 2021 Sep;9(3):758-770. doi: 10.1002/iid3.430. Epub 2021 May 7. Immun Inflamm Dis. 2021. PMID: 33960699 Free PMC article.
-
BAFF knockout improves systemic inflammation via regulating adipose tissue distribution in high-fat diet-induced obesity.Exp Mol Med. 2015 Jan 16;47(1):e129. doi: 10.1038/emm.2014.98. Exp Mol Med. 2015. PMID: 25591987 Free PMC article.
-
Hyperactivated MyD88 signaling in dendritic cells, through specific deletion of Lyn kinase, causes severe autoimmunity and inflammation.Proc Natl Acad Sci U S A. 2013 Aug 27;110(35):E3311-20. doi: 10.1073/pnas.1300617110. Epub 2013 Aug 12. Proc Natl Acad Sci U S A. 2013. PMID: 23940344 Free PMC article.
-
Deletion of Syk in neutrophils prevents immune complex arthritis.J Immunol. 2011 Oct 15;187(8):4319-30. doi: 10.4049/jimmunol.1100341. Epub 2011 Sep 14. J Immunol. 2011. PMID: 21918195 Free PMC article.
-
The structural wedge domain of the receptor-like tyrosine phosphatase CD45 enforces B cell tolerance by regulating substrate specificity.J Immunol. 2013 Mar 15;190(6):2527-35. doi: 10.4049/jimmunol.1202928. Epub 2013 Feb 8. J Immunol. 2013. PMID: 23396948 Free PMC article.
References
-
- Cash H., Relle M., Menke J., Brochhausen C., Jones S.A., Topley N., Galle P.R., Schwarting A. 2010. Interleukin 6 (IL-6) deficiency delays lupus nephritis in MRL-Faslpr mice: the IL-6 pathway as a new therapeutic target in treatment of autoimmune kidney disease in systemic lupus erythematosus. J. Rheumatol. 37:60–70 10.3899/jrheum.090194 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
