

Spatiotemporal organization, regulation, and functions of tractions during neutrophil chemotaxis
- PMID: 20616216
- PMCID: PMC2995358
- DOI: 10.1182/blood-2009-12-260851
Spatiotemporal organization, regulation, and functions of tractions during neutrophil chemotaxis
Abstract
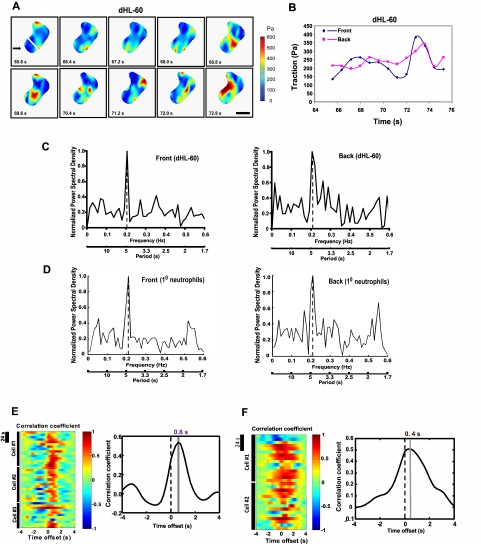
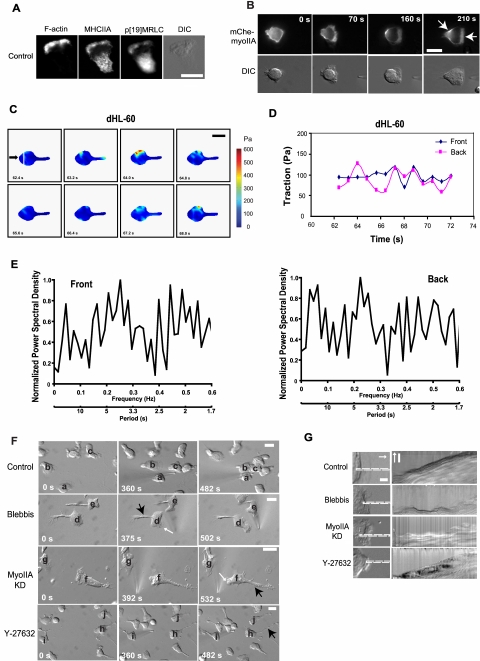
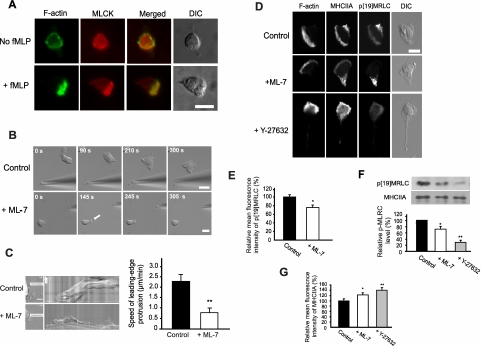
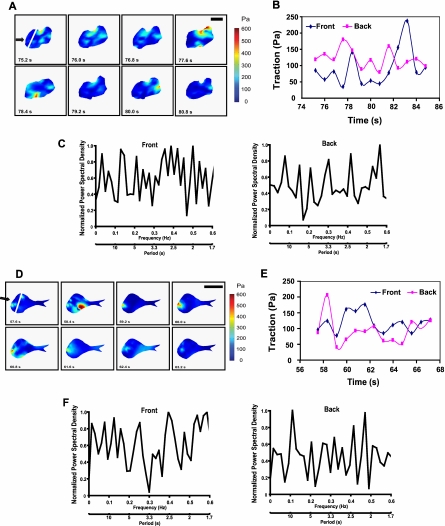
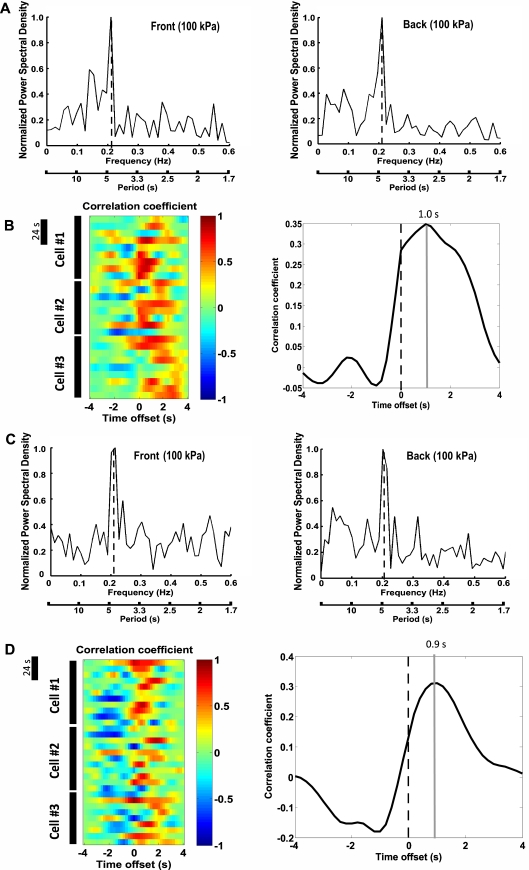
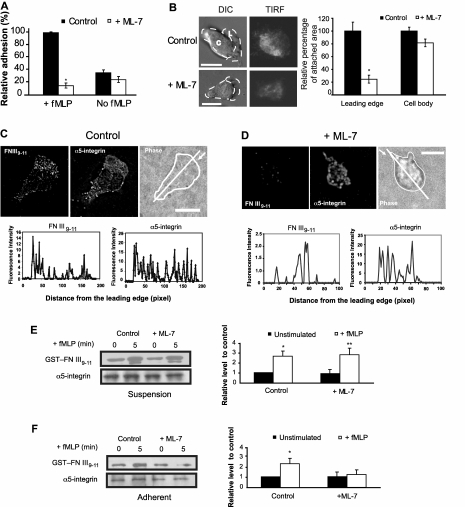
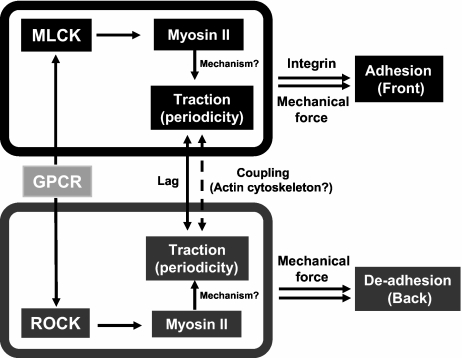
Despite recent advances in our understanding of biochemical regulation of neutrophil chemotaxis, little is known about how mechanical factors control neutrophils' persistent polarity and rapid motility. Here, using a human neutrophil-like cell line and human primary neutrophils, we describe a dynamic spatiotemporal pattern of tractions during chemotaxis. Tractions are located at both the leading and the trailing edge of neutrophils, where they oscillate with a defined periodicity. Interestingly, traction oscillations at the leading and the trailing edge are out of phase with the tractions at the front leading those at the back, suggesting a temporal mechanism that coordinates leading edge and trailing edge activities. The magnitude and periodicity of tractions depend on the activity of nonmuscle myosin IIA. Specifically, traction development at the leading edge requires myosin light chain kinase-mediated myosin II contractility and is necessary for α5β1-integrin activation and leading edge adhesion. Localized myosin II activation induced by spatially activated small GTPase Rho, and its downstream kinase p160-ROCK, as previously reported, leads to contraction of actin-myosin II complexes at the trailing edge, causing it to de-adhere. Our data identify a key biomechanical mechanism for persistent cell polarity and motility.
Figures
Similar articles
-
The non-receptor tyrosine kinase Lyn controls neutrophil adhesion by recruiting the CrkL-C3G complex and activating Rap1 at the leading edge.J Cell Sci. 2011 Jul 1;124(Pt 13):2153-64. doi: 10.1242/jcs.078535. Epub 2011 May 31. J Cell Sci. 2011. PMID: 21628423 Free PMC article.
-
Rac1 links leading edge and uropod events through Rho and myosin activation during chemotaxis.Blood. 2006 Oct 15;108(8):2814-20. doi: 10.1182/blood-2006-01-010363. Epub 2006 Jun 29. Blood. 2006. PMID: 16809619 Free PMC article.
-
Hem-1 complexes are essential for Rac activation, actin polymerization, and myosin regulation during neutrophil chemotaxis.PLoS Biol. 2006 Feb;4(2):e38. doi: 10.1371/journal.pbio.0040038. Epub 2006 Jan 24. PLoS Biol. 2006. PMID: 16417406 Free PMC article.
-
Signaling to migration in neutrophils: importance of localized pathways.Int J Biochem Cell Biol. 2003 Dec;35(12):1619-38. doi: 10.1016/s1357-2725(03)00144-4. Int J Biochem Cell Biol. 2003. PMID: 12962702 Review.
-
Signal transduction in neutrophil chemotaxis.Biochemistry (Mosc). 2001 Apr;66(4):351-68. doi: 10.1023/a:1010293809553. Biochemistry (Mosc). 2001. PMID: 11403641 Review.
Cited by
-
Reduced PaxillinB localization to cell-substrate adhesions promotes cell migration in Dictyostelium.bioRxiv [Preprint]. 2024 Mar 22:2024.03.19.585764. doi: 10.1101/2024.03.19.585764. bioRxiv. 2024. PMID: 38562712 Free PMC article. Preprint.
-
The non-receptor tyrosine kinase Lyn controls neutrophil adhesion by recruiting the CrkL-C3G complex and activating Rap1 at the leading edge.J Cell Sci. 2011 Jul 1;124(Pt 13):2153-64. doi: 10.1242/jcs.078535. Epub 2011 May 31. J Cell Sci. 2011. PMID: 21628423 Free PMC article.
-
Hybrid mechanosensing system to generate the polarity needed for migration in fish keratocytes.Cell Adh Migr. 2016 Jul 3;10(4):406-18. doi: 10.1080/19336918.2016.1170268. Epub 2016 Apr 28. Cell Adh Migr. 2016. PMID: 27124267 Free PMC article.
-
Helicobacter pylori-infected human neutrophils exhibit impaired chemotaxis and a uropod retraction defect.Front Immunol. 2022 Oct 20;13:1038349. doi: 10.3389/fimmu.2022.1038349. eCollection 2022. Front Immunol. 2022. PMID: 36341418 Free PMC article.
-
Both contractile axial and lateral traction force dynamics drive amoeboid cell motility.J Cell Biol. 2014 Mar 17;204(6):1045-61. doi: 10.1083/jcb.201307106. J Cell Biol. 2014. PMID: 24637328 Free PMC article.
References
-
- Wang F, Herzmark P, Weiner OD, Srinivasan S, Servant G, Bourne HR. Lipid products of PI(3)Ks maintain persistent cell polarity and directed motility in neutrophils. Nat Cell Biol. 2002;4(7):513–518. - PubMed
-
- Xu J, Wang F, Van Keymeulen A, et al. Divergent signals and cytoskeletal assemblies regulate self-organizing polarity in neutrophils. Cell. 2003;114(2):201–214. - PubMed
-
- Rottner K, Hall A, Small JV. Interplay between Rac and Rho in the control of substrate contact dynamics. Curr Biol. 1999;9(12):640–649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
