

Peripheral protein quality control removes unfolded CFTR from the plasma membrane
- PMID: 20595578
- PMCID: PMC5026491
- DOI: 10.1126/science.1191542
Peripheral protein quality control removes unfolded CFTR from the plasma membrane
Abstract
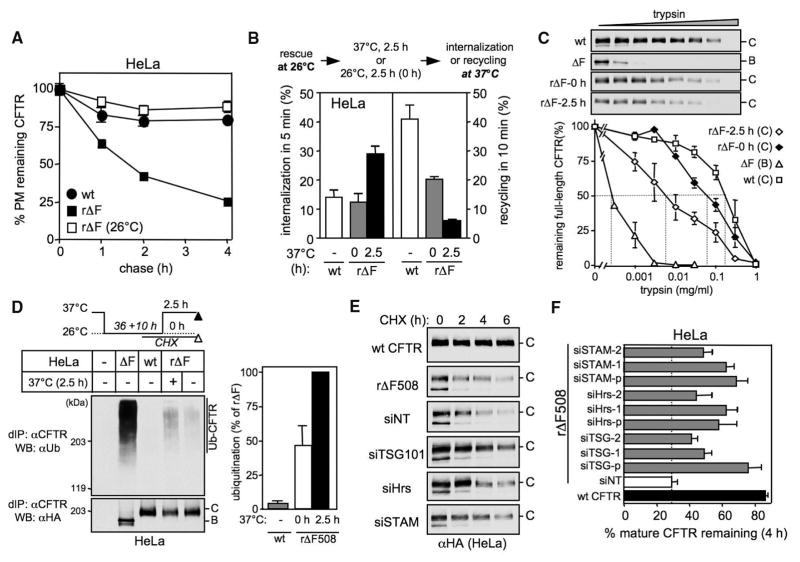
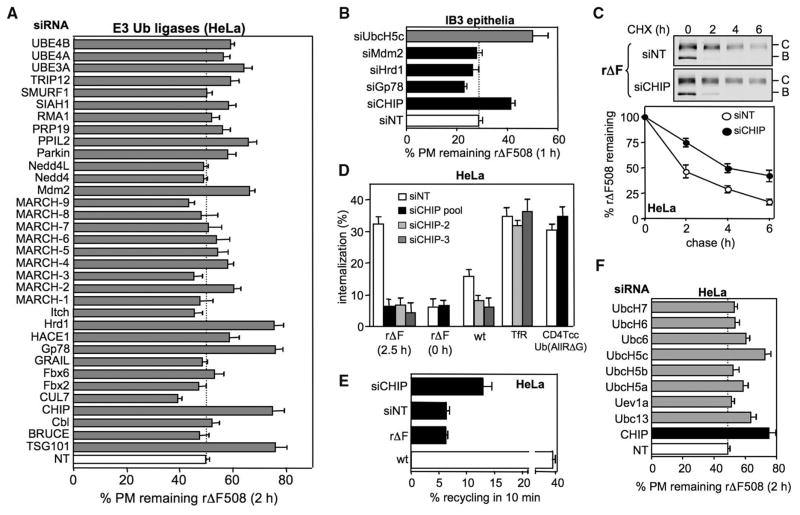
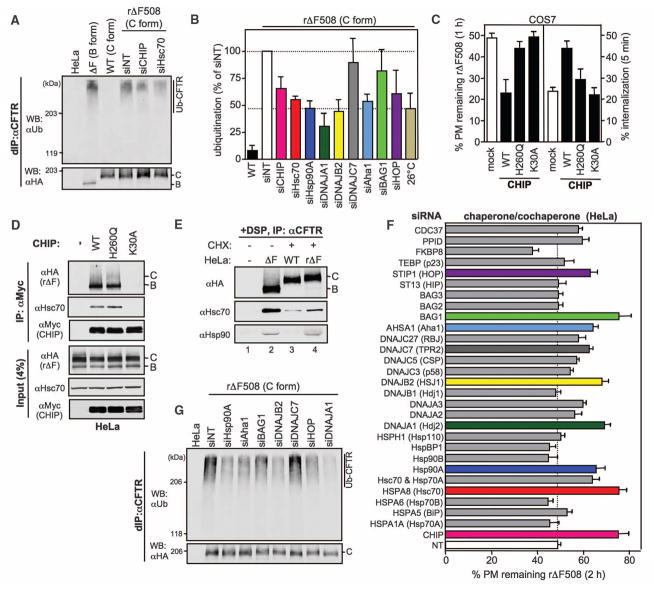
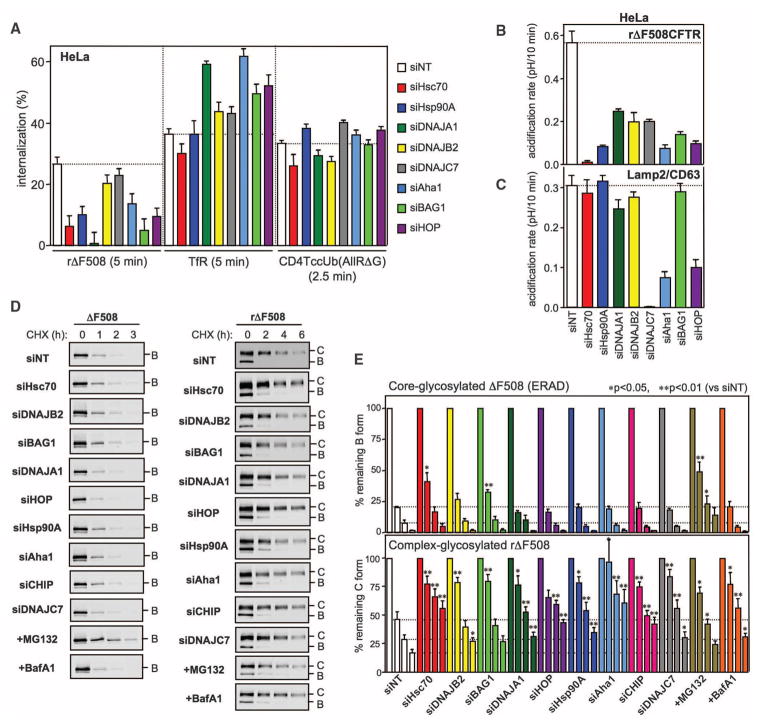
Therapeutic efforts to restore biosynthetic processing of the cystic fibrosis transmembrane conductance regulator lacking the F508 residue (DeltaF508CFTR) are hampered by ubiquitin-dependent lysosomal degradation of nonnative, rescued DeltaF508CFTR from the plasma membrane. Here, functional small interfering RNA screens revealed the contribution of chaperones, cochaperones, and ubiquitin-conjugating and -ligating enzymes to the elimination of unfolded CFTR from the cell surface, as part of a peripheral protein quality-control system. Ubiquitination of nonnative CFTR was required for efficient internalization and lysosomal degradation. This peripheral protein quality-control mechanism probably participates in the preservation of cellular homeostasis by degrading damaged plasma membrane proteins that have escaped from the endoplasmic reticulum quality control or are generated by environmental stresses in situ.
Figures

Comment in
-
Cell Biology. The proteome in balance.Science. 2010 Aug 13;329(5993):766-7. doi: 10.1126/science.1194160. Science. 2010. PMID: 20705837 Free PMC article. No abstract available.
Similar articles
-
Cell Biology. The proteome in balance.Science. 2010 Aug 13;329(5993):766-7. doi: 10.1126/science.1194160. Science. 2010. PMID: 20705837 Free PMC article. No abstract available.
-
Reduced PDZ interactions of rescued ΔF508CFTR increases its cell surface mobility.J Biol Chem. 2012 Dec 21;287(52):43630-8. doi: 10.1074/jbc.M112.421172. Epub 2012 Oct 31. J Biol Chem. 2012. PMID: 23115232 Free PMC article.
-
Chaperones rescue the energetic landscape of mutant CFTR at single molecule and in cell.Nat Commun. 2017 Aug 30;8(1):398. doi: 10.1038/s41467-017-00444-4. Nat Commun. 2017. PMID: 28855508 Free PMC article.
-
Cystic fibrosis transmembrane conductance regulator degradation: cross-talk between the ubiquitylation and SUMOylation pathways.FEBS J. 2013 Sep;280(18):4430-8. doi: 10.1111/febs.12415. Epub 2013 Jul 22. FEBS J. 2013. PMID: 23809253 Free PMC article. Review.
-
Regulation of CFTR Biogenesis by the Proteostatic Network and Pharmacological Modulators.Int J Mol Sci. 2020 Jan 10;21(2):452. doi: 10.3390/ijms21020452. Int J Mol Sci. 2020. PMID: 31936842 Free PMC article. Review.
Cited by
-
UBE3C Facilitates the ER-Associated and Peripheral Degradation of Misfolded CFTR.Cells. 2023 Nov 30;12(23):2741. doi: 10.3390/cells12232741. Cells. 2023. PMID: 38067172 Free PMC article.
-
Managing the underlying cause of cystic fibrosis: a future role for potentiators and correctors.Paediatr Drugs. 2013 Oct;15(5):393-402. doi: 10.1007/s40272-013-0035-3. Paediatr Drugs. 2013. PMID: 23757197 Review.
-
Phosphorylation-dependent 14-3-3 protein interactions regulate CFTR biogenesis.Mol Biol Cell. 2012 Mar;23(6):996-1009. doi: 10.1091/mbc.E11-08-0662. Epub 2012 Jan 25. Mol Biol Cell. 2012. PMID: 22278744 Free PMC article.
-
Calpain inhibition promotes the rescue of F(508)del-CFTR in PBMC from cystic fibrosis patients.PLoS One. 2013 Jun 13;8(6):e66089. doi: 10.1371/journal.pone.0066089. Print 2013. PLoS One. 2013. PMID: 23785472 Free PMC article.
-
Cystic fibrosis transmembrane conductance regulator (CFTR) potentiator VX-770 (ivacaftor) opens the defective channel gate of mutant CFTR in a phosphorylation-dependent but ATP-independent manner.J Biol Chem. 2012 Oct 26;287(44):36639-49. doi: 10.1074/jbc.M112.393637. Epub 2012 Aug 31. J Biol Chem. 2012. PMID: 22942289 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
