

Ex vivo cultures of microglia from young and aged rodent brain reveal age-related changes in microglial function
- PMID: 20580465
- PMCID: PMC4162517
- DOI: 10.1016/j.neurobiolaging.2010.05.008
Ex vivo cultures of microglia from young and aged rodent brain reveal age-related changes in microglial function
Abstract
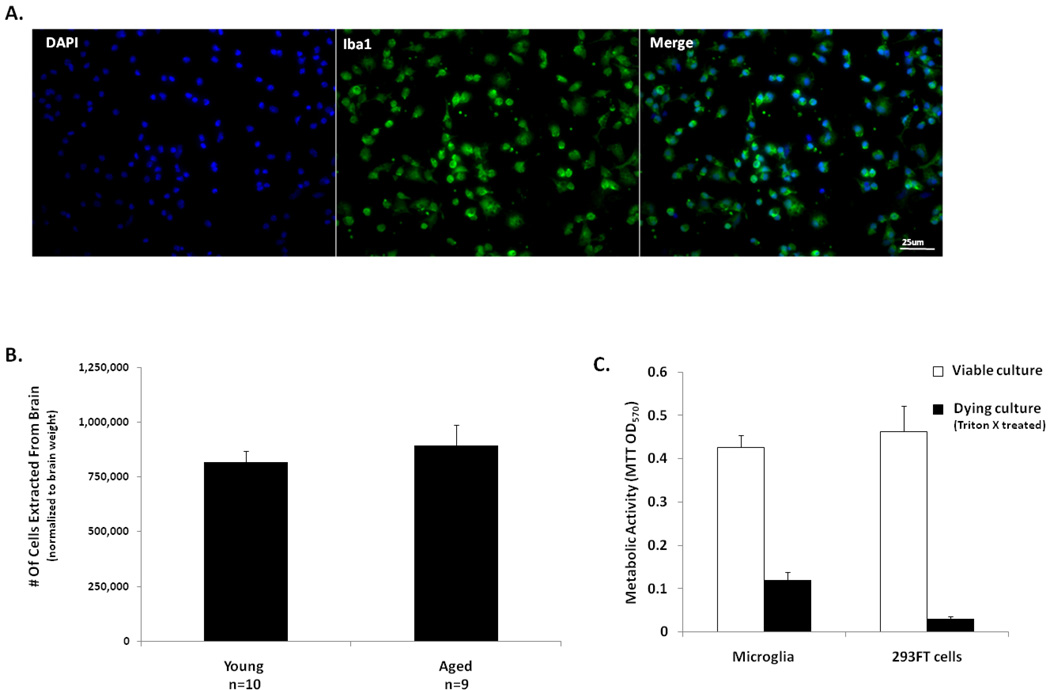
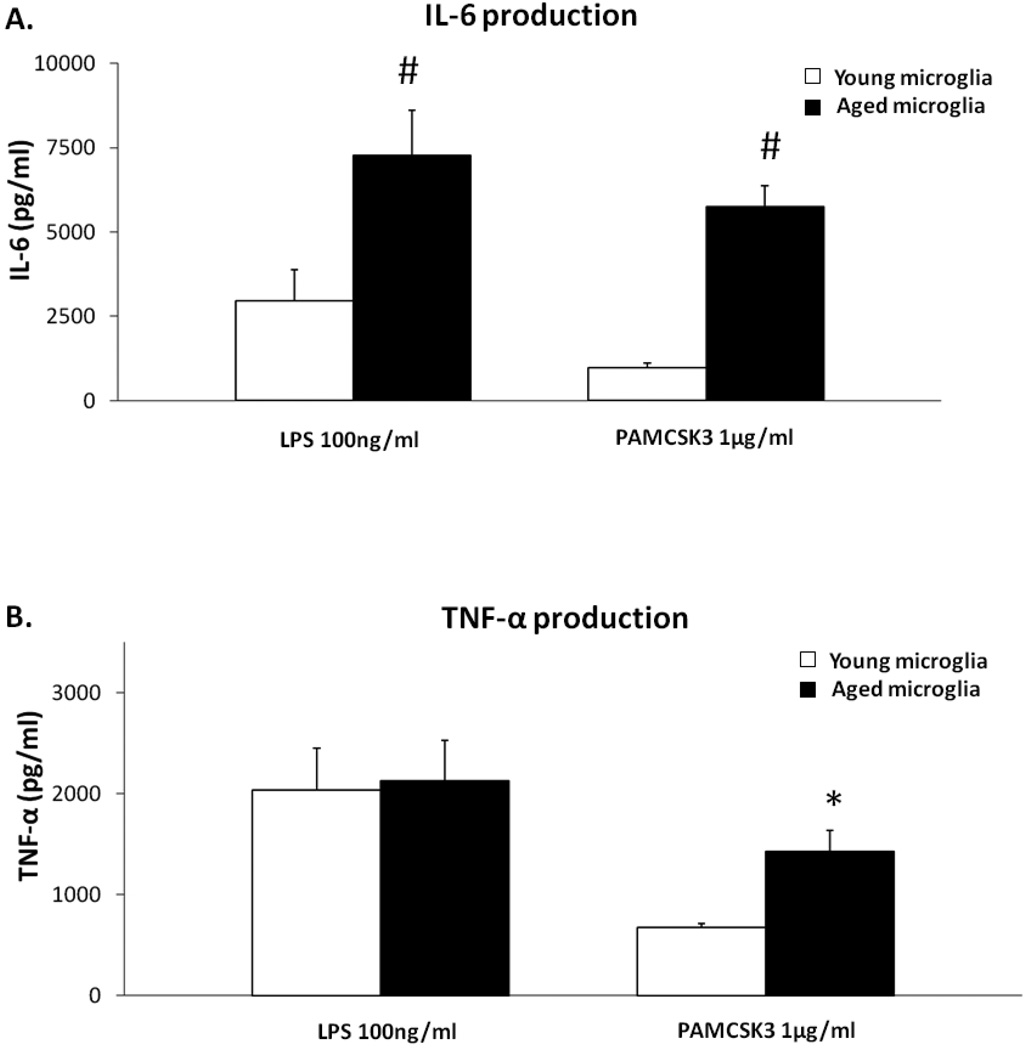
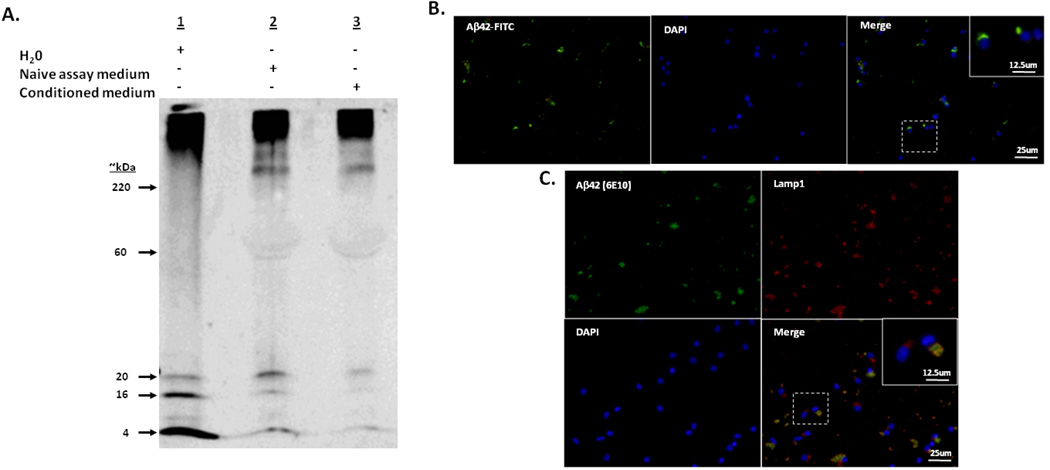
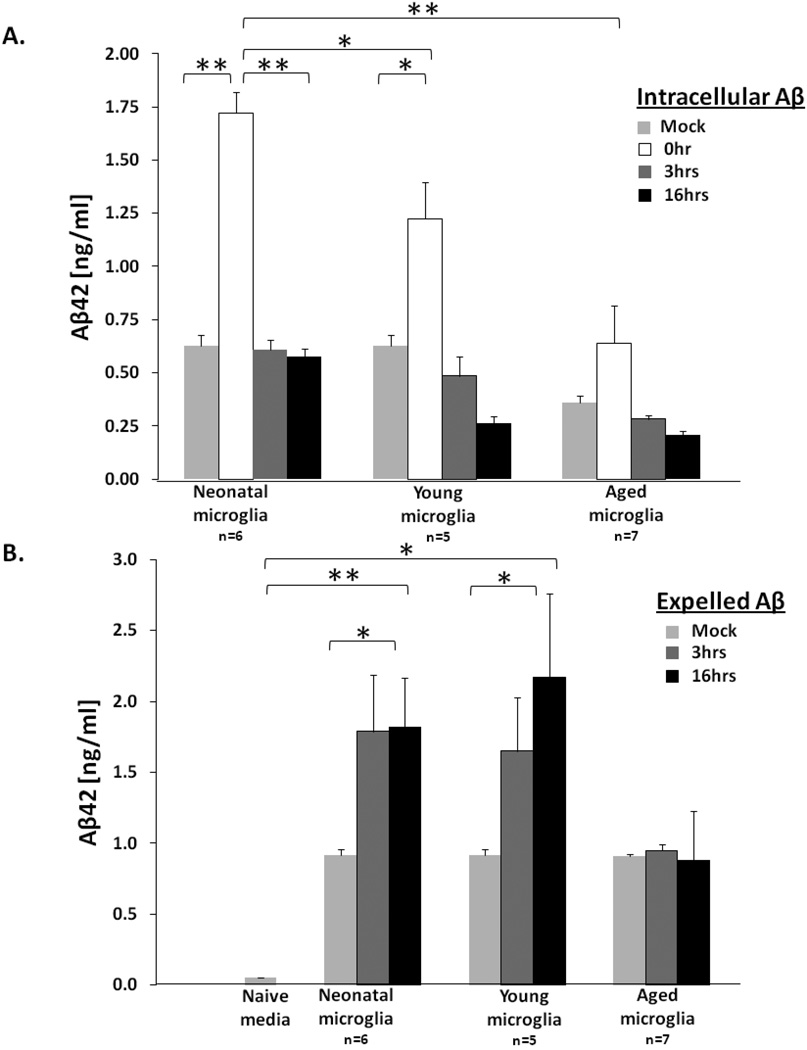
To understand how microglial cell function may change with aging, various protocols have been developed to isolate microglia from the young and aged central nervous system (CNS). Here we report modification of an existing protocol that is marked by less debris contamination and improved yields and demonstrate that microglial functions are varied and dependent on age. Specifically, we found that microglia from aged mice constitutively secrete greater amounts of interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) relative to microglia from younger mice and are less responsive to stimulation. Also, microglia from aged mice have reduced glutathione levels and internalize less amyloid beta peptide (Aβ) while microglia from mice of all ages do not retain the amyloid beta peptide for a significant length of time. These studies offer further support for the idea that microglial cell function changes with aging. They suggest that microglial Aβ phagocytosis results in Aβ redistribution rather than biophysical degradation in vivo and thereby provide mechanistic insight to the lack of amyloid burden elimination by parenchymal microglia in aged adults and those suffering from Alzheimer's disease.
Copyright © 2012. Published by Elsevier Inc.
Conflict of interest statement
None.
Figures

Similar articles
-
Role of Suppressor of Cytokine Signaling 3 (SOCS3) in Altering Activated Microglia Phenotype in APPswe/PS1dE9 Mice.J Alzheimers Dis. 2017;55(3):1235-1247. doi: 10.3233/JAD-160887. J Alzheimers Dis. 2017. PMID: 27814300
-
Microglial dysfunction and defective beta-amyloid clearance pathways in aging Alzheimer's disease mice.J Neurosci. 2008 Aug 13;28(33):8354-60. doi: 10.1523/JNEUROSCI.0616-08.2008. J Neurosci. 2008. PMID: 18701698 Free PMC article.
-
CD45 opposes beta-amyloid peptide-induced microglial activation via inhibition of p44/42 mitogen-activated protein kinase.J Neurosci. 2000 Oct 15;20(20):7587-94. doi: 10.1523/JNEUROSCI.20-20-07587.2000. J Neurosci. 2000. PMID: 11027218 Free PMC article.
-
The endocytic pathway in microglia during health, aging and Alzheimer's disease.Ageing Res Rev. 2016 Dec;32:89-103. doi: 10.1016/j.arr.2016.07.002. Epub 2016 Jul 12. Ageing Res Rev. 2016. PMID: 27421577 Free PMC article. Review.
-
Modeling microglial activation in Alzheimer's disease with human postmortem microglial cultures.Neurobiol Aging. 2001 Nov-Dec;22(6):945-56. doi: 10.1016/s0197-4580(01)00311-6. Neurobiol Aging. 2001. PMID: 11755003 Review.
Cited by
-
Clearance of amyloid-β peptides by microglia and macrophages: the issue of what, when and where.Future Neurol. 2012 Mar 1;7(2):165-176. doi: 10.2217/fnl.12.6. Future Neurol. 2012. PMID: 22737039 Free PMC article.
-
Aging-induced Nrf2-ARE pathway disruption in the subventricular zone drives neurogenic impairment in parkinsonian mice via PI3K-Wnt/β-catenin dysregulation.J Neurosci. 2013 Jan 23;33(4):1462-85. doi: 10.1523/JNEUROSCI.3206-12.2013. J Neurosci. 2013. PMID: 23345222 Free PMC article.
-
Glia-Derived Extracellular Vesicles in Parkinson's Disease.J Clin Med. 2020 Jun 21;9(6):1941. doi: 10.3390/jcm9061941. J Clin Med. 2020. PMID: 32575923 Free PMC article. Review.
-
Interplay Between Microglia and Alzheimer's Disease-Focus on the Most Relevant Risks: APOE Genotype, Sex and Age.Front Aging Neurosci. 2021 Apr 8;13:631827. doi: 10.3389/fnagi.2021.631827. eCollection 2021. Front Aging Neurosci. 2021. PMID: 33897406 Free PMC article.
-
Cannabidiol as an emergent therapeutic strategy for lessening the impact of inflammation on oxidative stress.Free Radic Biol Med. 2011 Sep 1;51(5):1054-61. doi: 10.1016/j.freeradbiomed.2011.01.007. Epub 2011 Jan 14. Free Radic Biol Med. 2011. PMID: 21238581 Free PMC article. Review.
References
-
- Boillee S, Yamanaka K, Lobsiger CS, Copeland NG, Jenkins NA, Kassiotis G, Kollias G, Cleveland DW. Onset and progression in inherited ALS determined by motor neurons and microglia. Science. 2006;312(5778):1389–1392. - PubMed
-
- Borchelt DR, Taraboulos A, Prusiner SB. Evidence for synthesis of scrapie prion proteins in the endocytic pathway. J Biol Chem. 1992;267(23):16188–16199. - PubMed
-
- Bouras C, Hof PR, Giannakopoulos P, Michel JP, Morrison JH. Regional distribution of neurofibrillary tangles and senile plaques in the cerebral cortex of elderly patients: a quantitative evaluation of a one-year autopsy population from a geriatric hospital. Cereb Cortex. 1994;4(2):138–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
