

The organization of local and distant functional connectivity in the human brain
- PMID: 20548945
- PMCID: PMC2883589
- DOI: 10.1371/journal.pcbi.1000808
The organization of local and distant functional connectivity in the human brain
Abstract
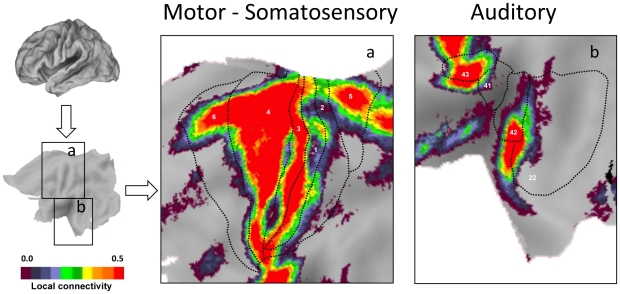
Information processing in the human brain arises from both interactions between adjacent areas and from distant projections that form distributed brain systems. Here we map interactions across different spatial scales by estimating the degree of intrinsic functional connectivity for the local (<or=14 mm) neighborhood directly surrounding brain regions as contrasted with distant (>14 mm) interactions. The balance between local and distant functional interactions measured at rest forms a map that separates sensorimotor cortices from heteromodal association areas and further identifies regions that possess both high local and distant cortical-cortical interactions. Map estimates of network measures demonstrate that high local connectivity is most often associated with a high clustering coefficient, long path length, and low physical cost. Task performance changed the balance between local and distant functional coupling in a subset of regions, particularly, increasing local functional coupling in regions engaged by the task. The observed properties suggest that the brain has evolved a balance that optimizes information-processing efficiency across different classes of specialized areas as well as mechanisms to modulate coupling in support of dynamically changing processing demands. We discuss the implications of these observations and applications of the present method for exploring normal and atypical brain function.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








Similar articles
-
Functional connectivity of distant cortical regions: role of remote synchronization and symmetry in interactions.Neuroimage. 2014 Aug 15;97:1-8. doi: 10.1016/j.neuroimage.2014.04.039. Epub 2014 Apr 24. Neuroimage. 2014. PMID: 24769266
-
Gradients of connectivity distance are anchored in primary cortex.Brain Struct Funct. 2017 Jul;222(5):2173-2182. doi: 10.1007/s00429-016-1333-7. Epub 2016 Nov 2. Brain Struct Funct. 2017. PMID: 27807628 Free PMC article.
-
Catecholaminergic Neuromodulation Shapes Intrinsic MRI Functional Connectivity in the Human Brain.J Neurosci. 2016 Jul 27;36(30):7865-76. doi: 10.1523/JNEUROSCI.0744-16.2016. J Neurosci. 2016. PMID: 27466332 Free PMC article.
-
Integrated technology for evaluation of brain function and neural plasticity.Phys Med Rehabil Clin N Am. 2004 Feb;15(1):263-306. doi: 10.1016/s1047-9651(03)00124-4. Phys Med Rehabil Clin N Am. 2004. PMID: 15029909 Review.
-
Clustered organization of cortical connectivity.Neuroinformatics. 2004;2(3):353-60. doi: 10.1385/NI:2:3:353. Neuroinformatics. 2004. PMID: 15365196 Review.
Cited by
-
The cross-sectional interplay between neurochemical profile and brain connectivity.Hum Brain Mapp. 2021 Jun 15;42(9):2722-2733. doi: 10.1002/hbm.25396. Epub 2021 Apr 9. Hum Brain Mapp. 2021. PMID: 33835605 Free PMC article.
-
Synchronization, non-linear dynamics and low-frequency fluctuations: analogy between spontaneous brain activity and networked single-transistor chaotic oscillators.Chaos. 2015 Mar;25(3):033107. doi: 10.1063/1.4914938. Chaos. 2015. PMID: 25833429 Free PMC article.
-
Test-retest reliability of graph metrics in high-resolution functional connectomics: a resting-state functional MRI study.CNS Neurosci Ther. 2015 Oct;21(10):802-16. doi: 10.1111/cns.12431. Epub 2015 Jul 27. CNS Neurosci Ther. 2015. PMID: 26212146 Free PMC article.
-
Social Brain Network of Children with Autism Spectrum Disorder: Characterization of Functional Connectivity and Potential Association with Stereotyped Behavior.Brain Sci. 2023 Feb 7;13(2):280. doi: 10.3390/brainsci13020280. Brain Sci. 2023. PMID: 36831823 Free PMC article.
-
Task effects on functional connectivity measures after stroke.Exp Brain Res. 2022 Feb;240(2):575-590. doi: 10.1007/s00221-021-06261-y. Epub 2021 Dec 2. Exp Brain Res. 2022. PMID: 34860257
References
-
- Felleman DJ, Van Essen DC. Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex. 1991;1:1–47. - PubMed
-
- Distler C, Boussaoud D, Desimone R, Ungerleider LG. Cortical connections of inferior temporal area TEO in macaque monkeys. J Comp Neurol. 1993;334:125–150. - PubMed
-
- Ungerleider LG, Haxby JV. ‘What’ and ‘where’ in the human brain. Curr Opin Neurobiol. 1994;4:157–165. - PubMed
-
- Jones EG, Powell TP. An anatomical study of converging sensory pathways within the cerebral cortex of the monkey. Brain. 1970;93:793–820. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
