

Induced fit, conformational selection and independent dynamic segments: an extended view of binding events
- PMID: 20541943
- PMCID: PMC3018770
- DOI: 10.1016/j.tibs.2010.04.009
Induced fit, conformational selection and independent dynamic segments: an extended view of binding events
Abstract
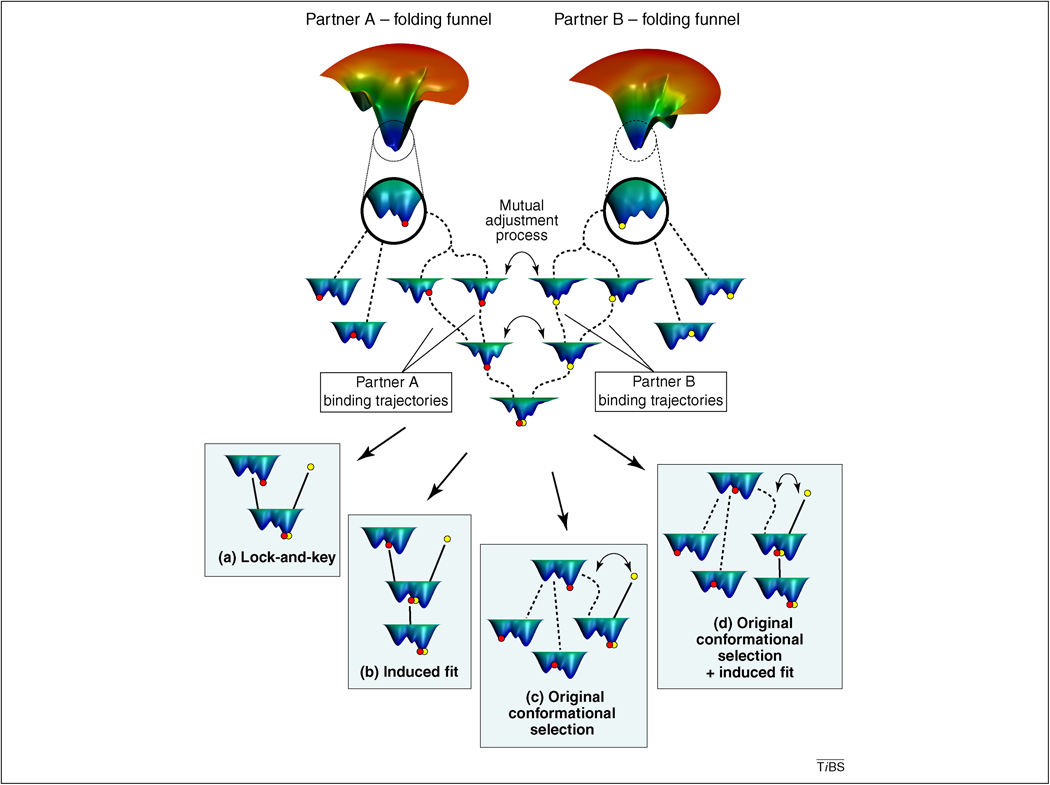
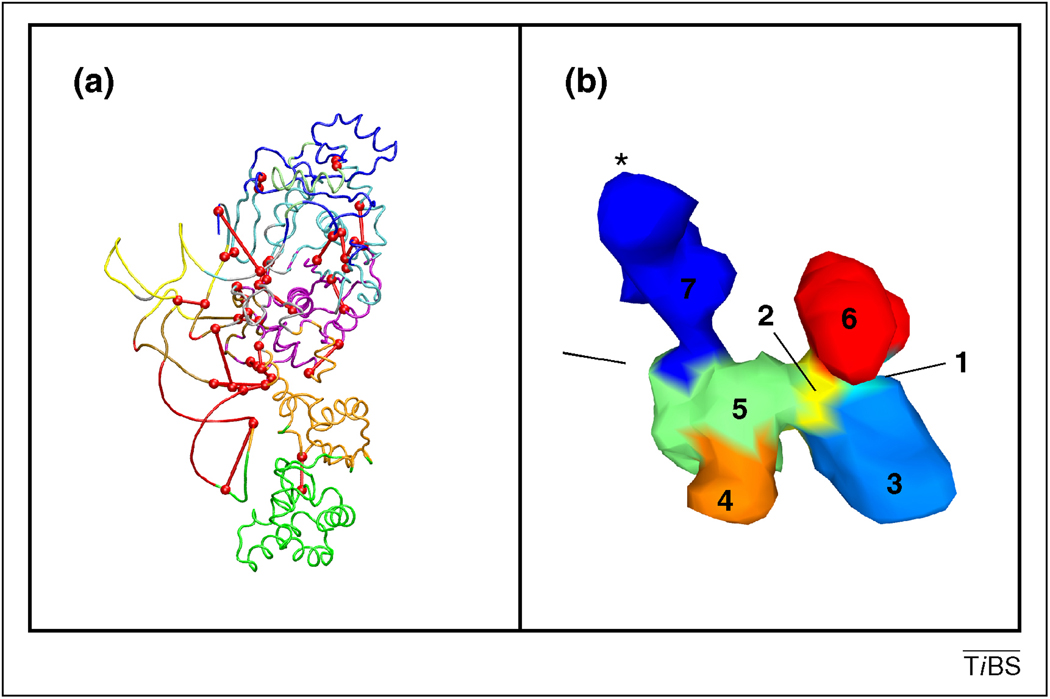
Single molecule and NMR measurements of protein dynamics increasingly uncover the complexity of binding scenarios. Here, we describe an extended conformational selection model that embraces a repertoire of selection and adjustment processes. Induced fit can be viewed as a subset of this repertoire, whose contribution is affected by the bond types stabilizing the interaction and the differences between the interacting partners. We argue that protein segments whose dynamics are distinct from the rest of the protein ('discrete breathers') can govern conformational transitions and allosteric propagation that accompany binding processes and, as such, might be more sensitive to mutational events. Additionally, we highlight the dynamic complexity of binding scenarios as they relate to events such as aggregation and signalling, and the crowded cellular environment.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Multiple conformational selection and induced fit events take place in allosteric propagation.Biophys Chem. 2014 Feb;186:22-30. doi: 10.1016/j.bpc.2013.10.002. Epub 2013 Oct 31. Biophys Chem. 2014. PMID: 24239303 Free PMC article. Review.
-
Macromolecular crowding: chemistry and physics meet biology (Ascona, Switzerland, 10-14 June 2012).Phys Biol. 2013 Aug;10(4):040301. doi: 10.1088/1478-3975/10/4/040301. Epub 2013 Aug 2. Phys Biol. 2013. PMID: 23912807
-
Conformational selection in protein binding and function.Protein Sci. 2014 Nov;23(11):1508-18. doi: 10.1002/pro.2539. Epub 2014 Sep 6. Protein Sci. 2014. PMID: 25155241 Free PMC article. Review.
-
Protein dynamics and conformational disorder in molecular recognition.J Mol Recognit. 2010 Mar-Apr;23(2):105-16. doi: 10.1002/jmr.961. J Mol Recognit. 2010. PMID: 19585546 Review.
-
Single molecule insights on conformational selection and induced fit mechanism.Biophys Chem. 2014 Feb;186:46-54. doi: 10.1016/j.bpc.2013.11.003. Epub 2013 Nov 13. Biophys Chem. 2014. PMID: 24342874 Review.
Cited by
-
Investigation of a Catenane with a Responsive Noncovalent Network: Mimicking Long-Range Responses in Proteins.J Am Chem Soc. 2016 Oct 12;138(40):13344-13352. doi: 10.1021/jacs.6b07833. Epub 2016 Sep 29. J Am Chem Soc. 2016. PMID: 27631725 Free PMC article.
-
Allostery and binding cooperativity of the catalytic subunit of protein kinase A by NMR spectroscopy and molecular dynamics simulations.Adv Protein Chem Struct Biol. 2012;87:363-89. doi: 10.1016/B978-0-12-398312-1.00012-3. Adv Protein Chem Struct Biol. 2012. PMID: 22607761 Free PMC article. Review.
-
Connecting the kinetics and energy landscape of tRNA translocation on the ribosome.PLoS Comput Biol. 2013;9(3):e1003003. doi: 10.1371/journal.pcbi.1003003. Epub 2013 Mar 21. PLoS Comput Biol. 2013. PMID: 23555233 Free PMC article.
-
Structured crowding and its effects on enzyme catalysis.Top Curr Chem. 2013;337:123-37. doi: 10.1007/128_2012_316. Top Curr Chem. 2013. PMID: 23571857 Free PMC article. Review.
-
A Database of Force-Field Parameters, Dynamics, and Properties of Antimicrobial Compounds.Molecules. 2015 Aug 3;20(8):13997-4021. doi: 10.3390/molecules200813997. Molecules. 2015. PMID: 26247924 Free PMC article.
References
-
- Fischer E. Einfluss der Configuration auf die Wirkung der Enzyme. Ber. Dtsch. Chem. Ges. 1894;27:2984–2993.
-
- Straub FB, Szabolcsi G. O dinamicseszkij aszpektah sztukturü fermentov. (On the dynamic aspects of protein structure) In: Braunstein AE, Russian, editors. Molecular Biology, Problems and Perspectives. Nauka, Moscow: Izdat; 1964. pp. 182–187.
-
- Závodszky P, Abaturov LV, Varshavsky YM. Structure of glyceraldehyde-3-phosphate dehydrogenase and its alteration by coenzyme binding. Acta Biochim. Biophys. Acad. Sci. Hung. 1966;1:389–403.
-
- Frauenfelder H, Sligar SG, Wolynes PG. The energy landscapes and motions of proteins. Science. 1991;254:1598–1603. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
