

Lysosomal proteolysis and autophagy require presenilin 1 and are disrupted by Alzheimer-related PS1 mutations
- PMID: 20541250
- PMCID: PMC3647462
- DOI: 10.1016/j.cell.2010.05.008
Lysosomal proteolysis and autophagy require presenilin 1 and are disrupted by Alzheimer-related PS1 mutations
Abstract
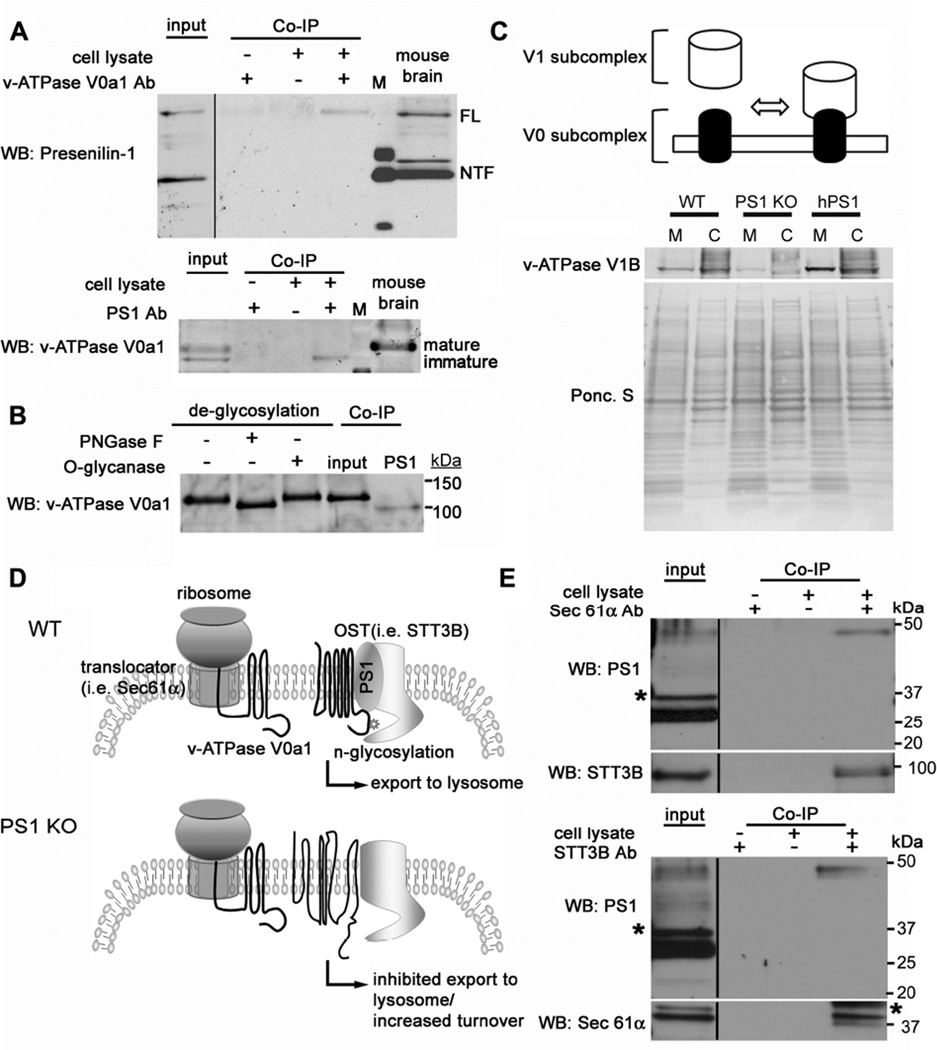
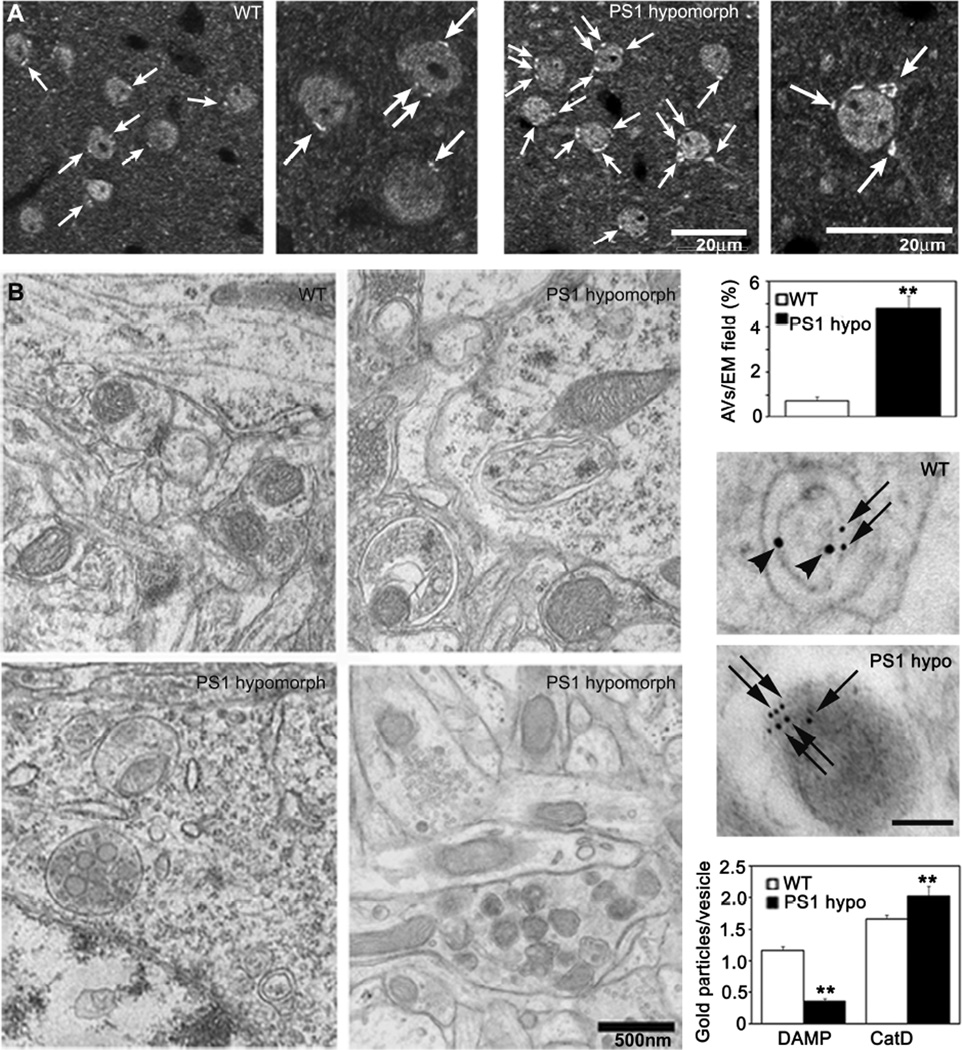
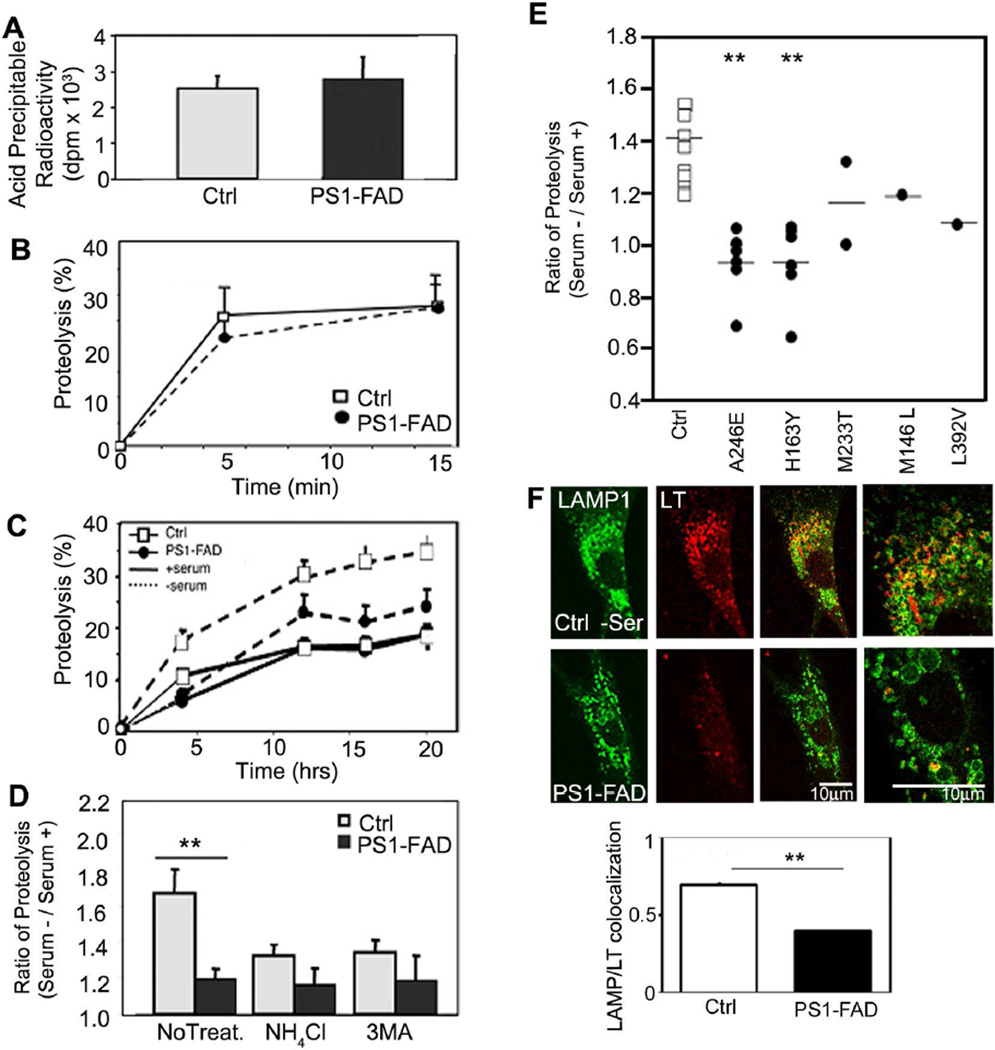
Macroautophagy is a lysosomal degradative pathway essential for neuron survival. Here, we show that macroautophagy requires the Alzheimer's disease (AD)-related protein presenilin-1 (PS1). In PS1 null blastocysts, neurons from mice hypomorphic for PS1 or conditionally depleted of PS1, substrate proteolysis and autophagosome clearance during macroautophagy are prevented as a result of a selective impairment of autolysosome acidification and cathepsin activation. These deficits are caused by failed PS1-dependent targeting of the v-ATPase V0a1 subunit to lysosomes. N-glycosylation of the V0a1 subunit, essential for its efficient ER-to-lysosome delivery, requires the selective binding of PS1 holoprotein to the unglycosylated subunit and the Sec61alpha/oligosaccharyltransferase complex. PS1 mutations causing early-onset AD produce a similar lysosomal/autophagy phenotype in fibroblasts from AD patients. PS1 is therefore essential for v-ATPase targeting to lysosomes, lysosome acidification, and proteolysis during autophagy. Defective lysosomal proteolysis represents a basis for pathogenic protein accumulations and neuronal cell death in AD and suggests previously unidentified therapeutic targets.
Figures
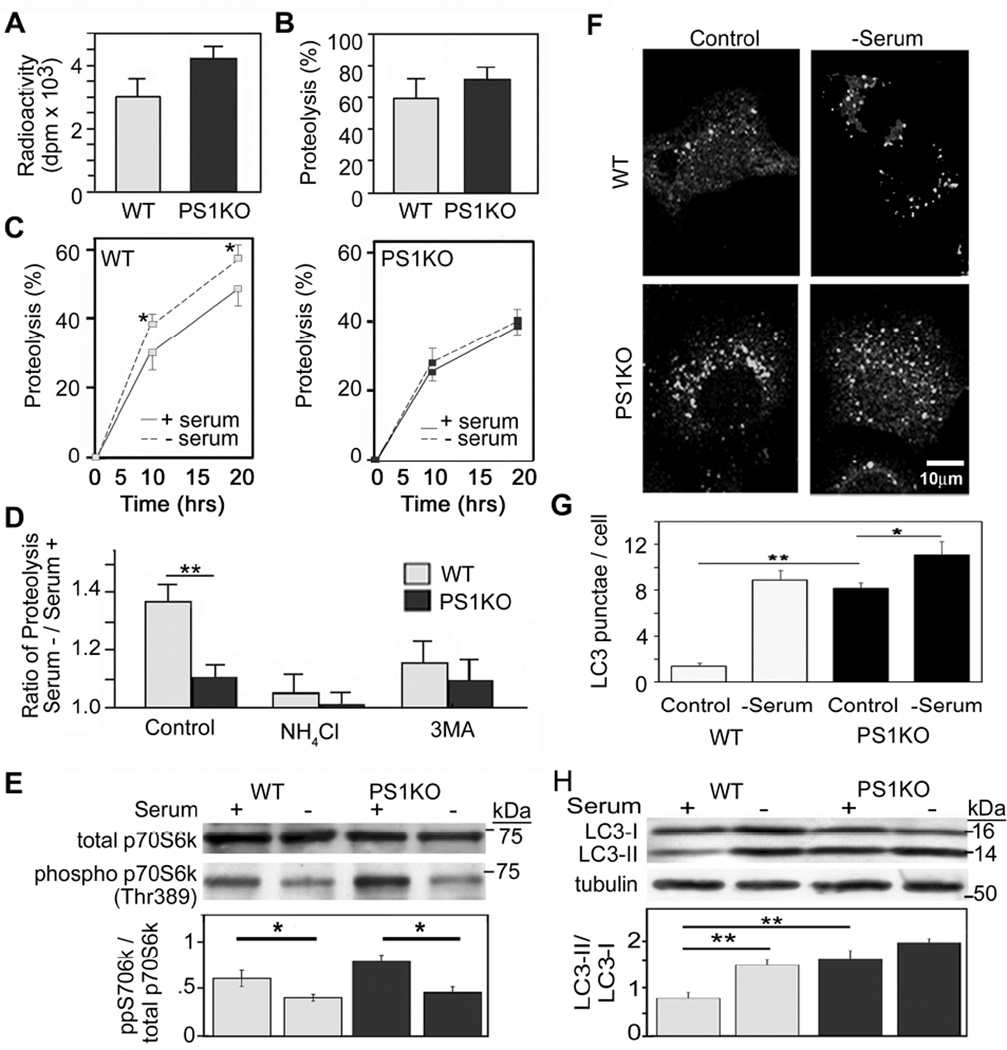
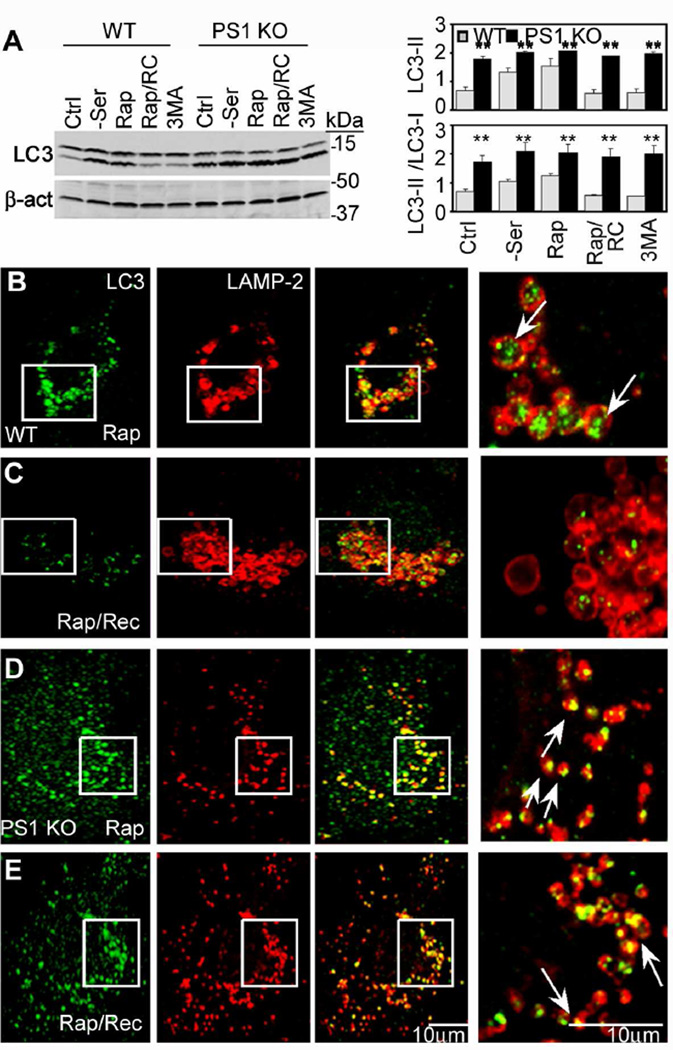
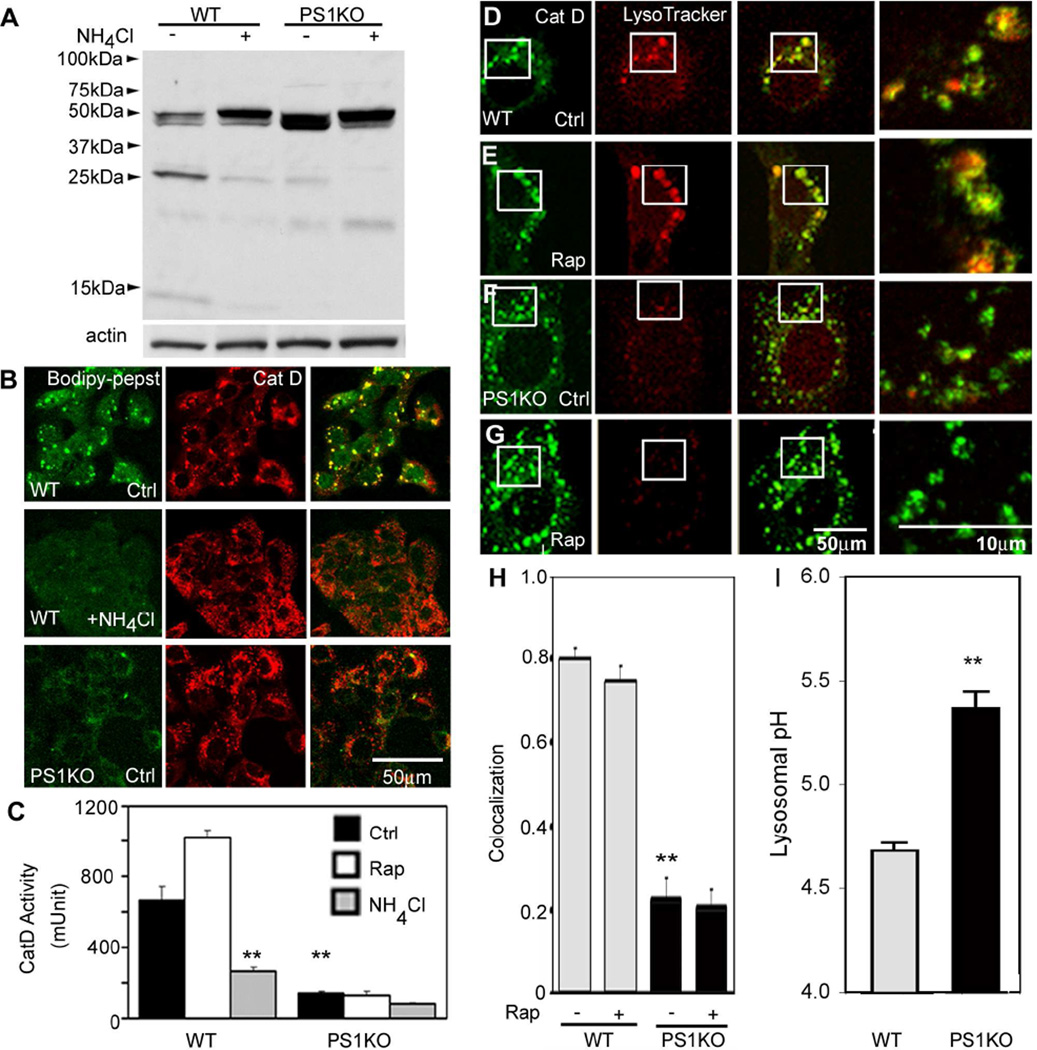
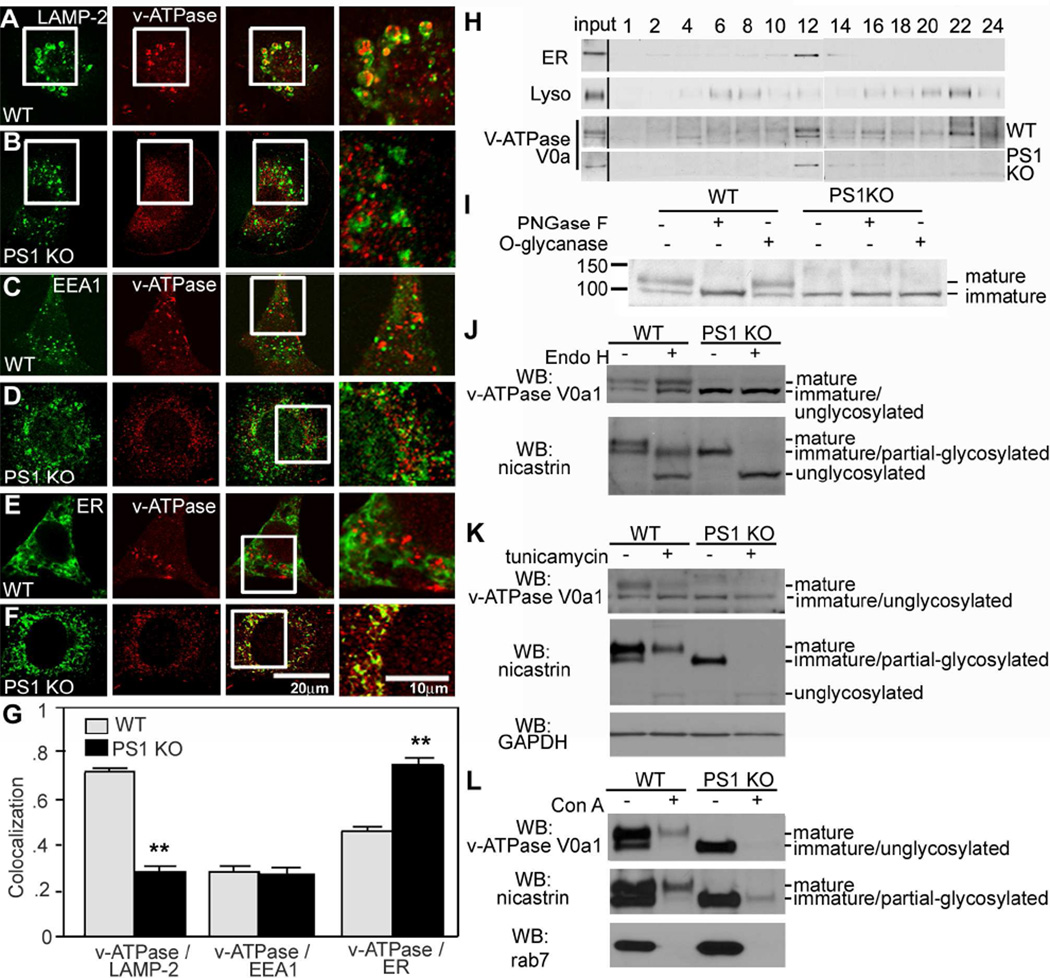
Comment in
-
Alzheimer's disease neurons fail the acid test.Cell. 2010 Jun 25;141(7):1112-4. doi: 10.1016/j.cell.2010.06.009. Cell. 2010. PMID: 20602994
Similar articles
-
Presenilin 1 Maintains Lysosomal Ca(2+) Homeostasis via TRPML1 by Regulating vATPase-Mediated Lysosome Acidification.Cell Rep. 2015 Sep 1;12(9):1430-44. doi: 10.1016/j.celrep.2015.07.050. Epub 2015 Aug 20. Cell Rep. 2015. PMID: 26299959 Free PMC article.
-
Autophagy failure in Alzheimer's disease and the role of defective lysosomal acidification.Eur J Neurosci. 2013 Jun;37(12):1949-61. doi: 10.1111/ejn.12169. Eur J Neurosci. 2013. PMID: 23773064 Free PMC article.
-
Lysosome and calcium dysregulation in Alzheimer's disease: partners in crime.Biochem Soc Trans. 2013 Dec;41(6):1495-502. doi: 10.1042/BST20130201. Biochem Soc Trans. 2013. PMID: 24256243 Free PMC article. Review.
-
A role for presenilins in autophagy revisited: normal acidification of lysosomes in cells lacking PSEN1 and PSEN2.J Neurosci. 2012 Jun 20;32(25):8633-48. doi: 10.1523/JNEUROSCI.0556-12.2012. J Neurosci. 2012. PMID: 22723704 Free PMC article.
-
The emerging roles of vacuolar-type ATPase-dependent Lysosomal acidification in neurodegenerative diseases.Transl Neurodegener. 2020 May 11;9(1):17. doi: 10.1186/s40035-020-00196-0. Transl Neurodegener. 2020. PMID: 32393395 Free PMC article. Review.
Cited by
-
Autophagosome dynamics in neurodegeneration at a glance.J Cell Sci. 2015 Apr 1;128(7):1259-67. doi: 10.1242/jcs.161216. J Cell Sci. 2015. PMID: 25829512 Free PMC article. Review.
-
Neuritic Plaques - Gateways to Understanding Alzheimer's Disease.Mol Neurobiol. 2024 May;61(5):2808-2821. doi: 10.1007/s12035-023-03736-7. Epub 2023 Nov 8. Mol Neurobiol. 2024. PMID: 37940777 Free PMC article. Review.
-
Recent advances in Alzheimer's disease: Mechanisms, clinical trials and new drug development strategies.Signal Transduct Target Ther. 2024 Aug 23;9(1):211. doi: 10.1038/s41392-024-01911-3. Signal Transduct Target Ther. 2024. PMID: 39174535 Free PMC article. Review.
-
Age-mediated transcriptomic changes in adult mouse substantia nigra.PLoS One. 2013 Apr 30;8(4):e62456. doi: 10.1371/journal.pone.0062456. Print 2013. PLoS One. 2013. PMID: 23638090 Free PMC article.
-
Axonal transport and neurodegenerative disease: can we see the elephant?Prog Neurobiol. 2012 Dec;99(3):186-90. doi: 10.1016/j.pneurobio.2012.03.006. Epub 2012 Apr 1. Prog Neurobiol. 2012. PMID: 22484448 Free PMC article. Review.
References
-
- Auteri JS, Okada A, Bochaki V, Dice JF. Regulation of intracellular protein degradation in IMR-90 human diploid fibroblasts. J Cell Physiol. 1983;115:167–174. - PubMed
-
- Bolt G, Kristensen C, Steenstrup TD. Posttranslational N-glycosylation takes place during the normal processing of human coagulation factor VII. Glycobiology. 2005;15:541–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
