

The T helper type 2 response to cysteine proteases requires dendritic cell-basophil cooperation via ROS-mediated signaling
- PMID: 20495560
- PMCID: PMC3145206
- DOI: 10.1038/ni.1883
The T helper type 2 response to cysteine proteases requires dendritic cell-basophil cooperation via ROS-mediated signaling
Abstract
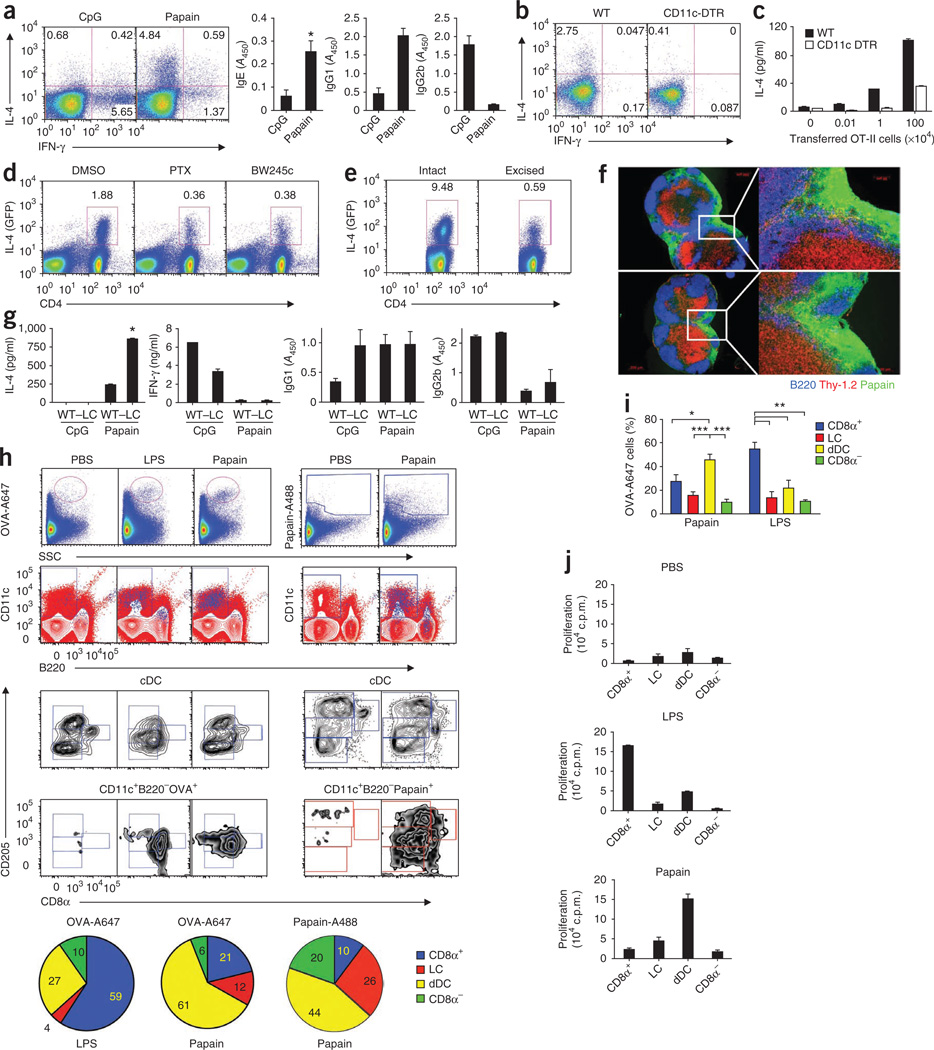
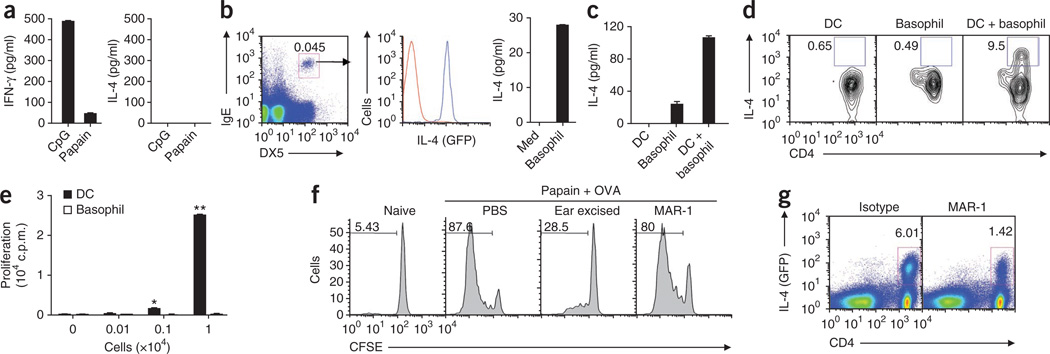
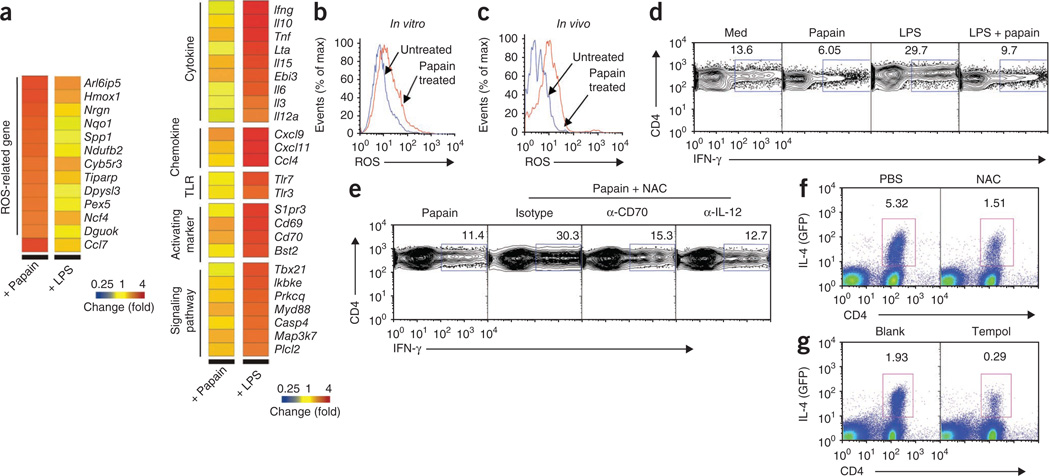
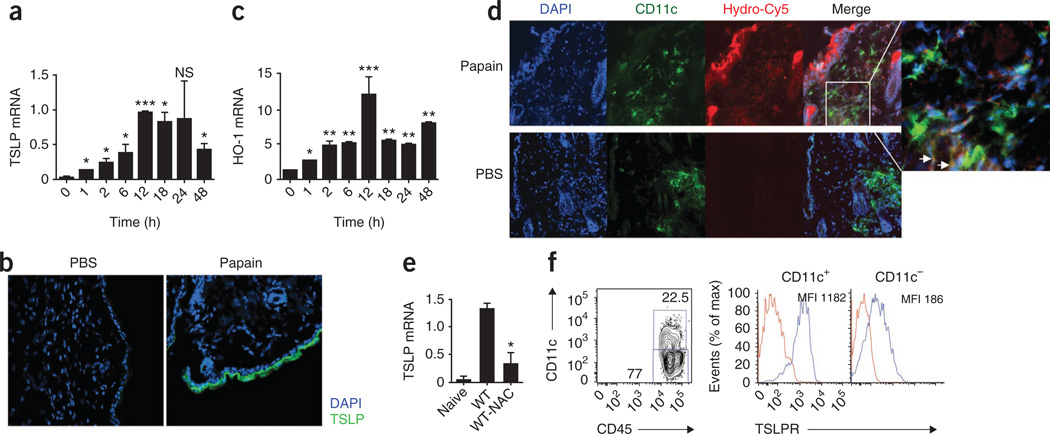
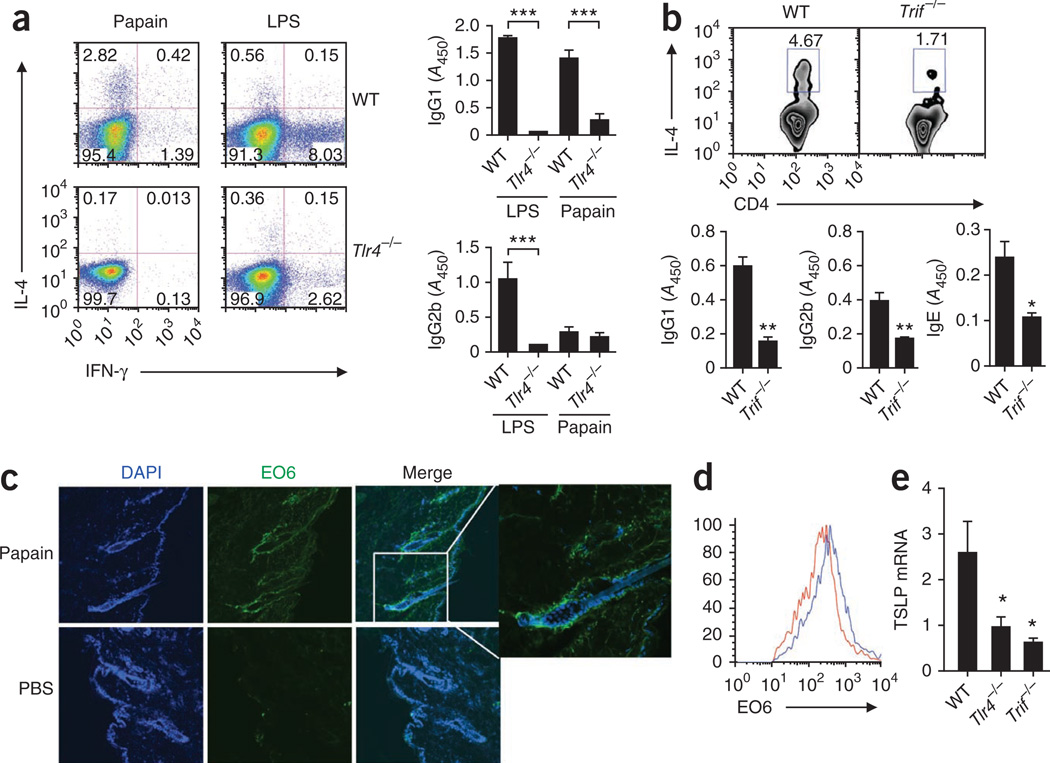
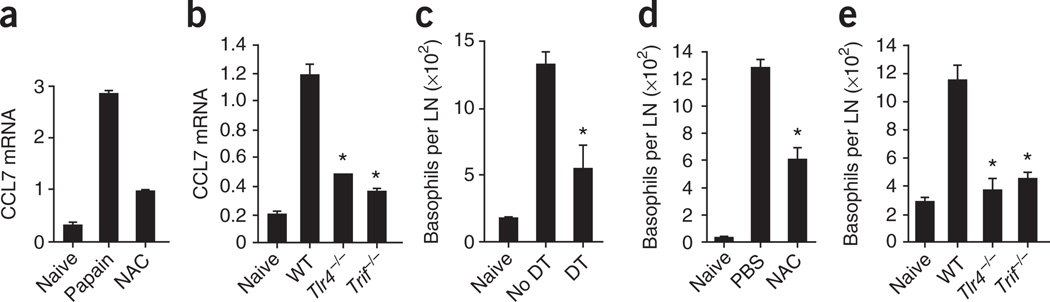
The mechanisms that initiate T helper type 2 (T(H)2) responses are poorly understood. Here we demonstrate that cysteine protease-induced T(H)2 responses occur via 'cooperation' between migratory dermal dendritic cells (DCs) and basophils positive for interleukin 4 (IL-4). Subcutaneous immunization with papain plus antigen induced reactive oxygen species (ROS) in lymph node DCs and in dermal DCs and epithelial cells of the skin. ROS orchestrated T(H)2 responses by inducing oxidized lipids that triggered the induction of thymic stromal lymphopoietin (TSLP) by epithelial cells mediated by Toll-like receptor 4 (TLR4) and the adaptor protein TRIF; by suppressing production of the T(H)1-inducing molecules IL-12 and CD70 in lymph node DCs; and by inducing the DC-derived chemokine CCL7, which mediated recruitment of IL-4(+) basophils to the lymph node. Thus, the T(H)2 response to cysteine proteases requires DC-basophil cooperation via ROS-mediated signaling.
Figures
Similar articles
-
Naive T cells sense the cysteine protease allergen papain through protease-activated receptor 2 and propel TH2 immunity.J Allergy Clin Immunol. 2012 May;129(5):1377-1386.e13. doi: 10.1016/j.jaci.2012.02.035. Epub 2012 Mar 27. J Allergy Clin Immunol. 2012. PMID: 22460072 Free PMC article.
-
Skin thymic stromal lymphopoietin initiates Th2 responses through an orchestrated immune cascade.Nat Commun. 2013;4:2847. doi: 10.1038/ncomms3847. Nat Commun. 2013. PMID: 24284909
-
Basophil-derived IL-4 promotes epicutaneous antigen sensitization concomitant with the development of food allergy.J Allergy Clin Immunol. 2018 Jan;141(1):223-234.e5. doi: 10.1016/j.jaci.2017.02.035. Epub 2017 Apr 6. J Allergy Clin Immunol. 2018. PMID: 28390860
-
Thymic stromal lymphopoietin, OX40-ligand, and interleukin-25 in allergic responses.Clin Exp Allergy. 2009 Jun;39(6):798-806. doi: 10.1111/j.1365-2222.2009.03241.x. Epub 2009 Apr 7. Clin Exp Allergy. 2009. PMID: 19400908 Free PMC article. Review.
-
Basophils induce Th2 immunity: is this final answer?Virulence. 2010 Sep-Oct;1(5):399-401. doi: 10.4161/viru.1.5.12550. Virulence. 2010. PMID: 21178477 Free PMC article. Review.
Cited by
-
Modulating Th2 Cell Immunity for the Treatment of Asthma.Front Immunol. 2021 Feb 10;12:637948. doi: 10.3389/fimmu.2021.637948. eCollection 2021. Front Immunol. 2021. PMID: 33643321 Free PMC article. Review.
-
TSLP promotes induction of Th2 differentiation but is not necessary during established allergen-induced pulmonary disease.PLoS One. 2013;8(2):e56433. doi: 10.1371/journal.pone.0056433. Epub 2013 Feb 20. PLoS One. 2013. PMID: 23437132 Free PMC article.
-
Functions of Murine Dendritic Cells.Immunity. 2016 Oct 18;45(4):719-736. doi: 10.1016/j.immuni.2016.10.010. Immunity. 2016. PMID: 27760337 Free PMC article. Review.
-
Naive T cells sense the cysteine protease allergen papain through protease-activated receptor 2 and propel TH2 immunity.J Allergy Clin Immunol. 2012 May;129(5):1377-1386.e13. doi: 10.1016/j.jaci.2012.02.035. Epub 2012 Mar 27. J Allergy Clin Immunol. 2012. PMID: 22460072 Free PMC article.
-
Chronic helminth infection reduces basophil responsiveness in an IL-10-dependent manner.J Immunol. 2012 May 1;188(9):4188-99. doi: 10.4049/jimmunol.1101859. Epub 2012 Mar 28. J Immunol. 2012. PMID: 22461700 Free PMC article.
References
-
- Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1989;7:145–173. - PubMed
-
- Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009;27:485–517. - PubMed
-
- Steinman RM, Banchereau J. Taking dendritic cells into medicine. Nature. 2007;449:419–426. - PubMed
-
- Steinman RM. Dendritic cells in vivo: a key target for a new vaccine science. Immunity. 2008;29:319–324. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R37 AI048638/AI/NIAID NIH HHS/United States
- HHSN266200700006C/AI/NIAID NIH HHS/United States
- R01 HL096796-03/HL/NHLBI NIH HHS/United States
- U54AI057157/AI/NIAID NIH HHS/United States
- R01 HL096796-02/HL/NHLBI NIH HHS/United States
- R01 HL096796/HL/NHLBI NIH HHS/United States
- R01 HL096796-01/HL/NHLBI NIH HHS/United States
- R37AI48638/AI/NIAID NIH HHS/United States
- N01 AI050019/AI/NIAID NIH HHS/United States
- U54 AI057157/AI/NIAID NIH HHS/United States
- HHSN266 200700006C/PHS HHS/United States
- R01DK057665/DK/NIDDK NIH HHS/United States
- U19 AI057266/AI/NIAID NIH HHS/United States
- N01 AI050025/AI/NIAID NIH HHS/United States
- R01 DK057665/DK/NIDDK NIH HHS/United States
- U19AI057266/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
