

Selectivity of the NF-{kappa}B response
- PMID: 20452937
- PMCID: PMC2845200
- DOI: 10.1101/cshperspect.a000257
Selectivity of the NF-{kappa}B response
Abstract
NF-kappaB is activated by many stimuli and NF-kappaB binding sites have been identified in a wide variety of genes. Yet, NF-kappaB-dependent gene expression must be stimulus- and cell-type-specific. In others words, the cellular response to different NF-kappaB activating stimuli, such as TNFalpha, IL-1, and LPS, must be different; and the response of different cell types, such as lymphocytes, fibroblasts, or epithelial cells, to the same NF-kappaB-inducing stimulus must also be different. Finally, kinetics of gene expression must be accounted for, so that all NF-kappaB-dependent genes are not activated simultaneously even if cell type and stimulus are constant. Here, we explore the mechanistic framework in which such regulatory aspects of NF-kappaB-dependent gene expression have been analyzed because they are likely to form the basis for physiological responses.
Figures
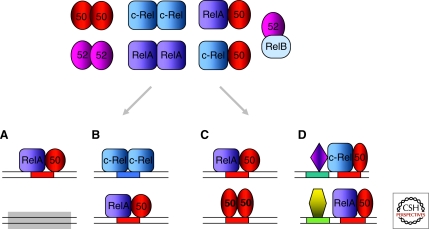
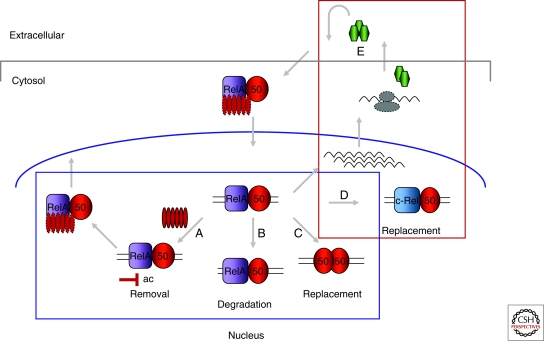
Similar articles
-
TNF/IL-1/NIK/NF-kappa B transduction pathway: a comparative study in normal and pathological human prostate (benign hyperplasia and carcinoma).Histopathology. 2008 Aug;53(2):166-76. doi: 10.1111/j.1365-2559.2008.03092.x. Histopathology. 2008. PMID: 18752500
-
Acute alcohol inhibits the induction of nuclear regulatory factor kappa B activation through CD14/toll-like receptor 4, interleukin-1, and tumor necrosis factor receptors: a common mechanism independent of inhibitory kappa B alpha degradation?Alcohol Clin Exp Res. 2002 Nov;26(11):1609-14. doi: 10.1097/01.ALC.0000036926.46632.57. Alcohol Clin Exp Res. 2002. PMID: 12436048
-
IkappaBalpha degradation and nuclear factor-kappaB DNA binding are insufficient for interleukin-1beta and tumor necrosis factor-alpha-induced kappaB-dependent transcription. Requirement for an additional activation pathway.J Biol Chem. 1998 Mar 20;273(12):6607-10. doi: 10.1074/jbc.273.12.6607. J Biol Chem. 1998. PMID: 9506955
-
Regulation and function of IKK and IKK-related kinases.Sci STKE. 2006 Oct 17;2006(357):re13. doi: 10.1126/stke.3572006re13. Sci STKE. 2006. PMID: 17047224 Review.
-
Identification of a nuclear factor kappa B-dependent gene network.Recent Prog Horm Res. 2003;58:95-130. doi: 10.1210/rp.58.1.95. Recent Prog Horm Res. 2003. PMID: 12795416 Review.
Cited by
-
Alternations of NF-κB Signaling by Natural Compounds in Muscle-Derived Cancers.Int J Mol Sci. 2023 Jul 25;24(15):11900. doi: 10.3390/ijms241511900. Int J Mol Sci. 2023. PMID: 37569275 Free PMC article. Review.
-
The Synergy of ADAM17-Induced Myocardial Inflammation and Metabolic Lipids Dysregulation During Acute Stress: New Pathophysiologic Insights Into Takotsubo Cardiomyopathy.Front Cardiovasc Med. 2021 Jun 4;8:696413. doi: 10.3389/fcvm.2021.696413. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34150874 Free PMC article. Review.
-
Discovery of RSV-Induced BRD4 Protein Interactions Using Native Immunoprecipitation and Parallel Accumulation-Serial Fragmentation (PASEF) Mass Spectrometry.Viruses. 2021 Mar 11;13(3):454. doi: 10.3390/v13030454. Viruses. 2021. PMID: 33799525 Free PMC article.
-
B cell TLRs and induction of immunoglobulin class-switch DNA recombination.Front Biosci (Landmark Ed). 2012 Jun 1;17(7):2594-615. doi: 10.2741/4073. Front Biosci (Landmark Ed). 2012. PMID: 22652800 Free PMC article. Review.
-
What Is Nuclear Factor Kappa B (NF-κB) Doing in and to the Mitochondrion?Front Cell Dev Biol. 2019 Aug 7;7:154. doi: 10.3389/fcell.2019.00154. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31448275 Free PMC article. Review.
References
-
- Banerjee D, Liou HC, Sen R 2005. c-Rel-dependent priming of naïve T cells by inflammatory cytokines. Immunity 4:445–458 - PubMed
-
- Bunting K, Rao S, Hardy K, Woltring D, Denyer GS, Wang J, Gerondakis S, Shannon MF 2007. Genome-wide analysis of gene expression in T cells to identify targets of the NF-κ B transcription factor c-Rel. J Immunol 178:7097–7109 - PubMed
-
- Carmody RJ, Ruan Q, Liou HC, Chen YH 2007. Essential roles of c-Rel in TLR-induced IL-23 19 gene expression in dendritic cells. J Immunol 178:186–191 - PubMed
-
- Chen FE, Ghosh G 1999. Regulation of DNA binding by Rel/NF-κB transcription factors: Structural views. Oncogene 18:6845–6852 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R21 AI076921-02/AI/NIAID NIH HHS/United States
- R01 CA127279/CA/NCI NIH HHS/United States
- R01 AI073868-05/AI/NIAID NIH HHS/United States
- R01 GM086372-04/GM/NIGMS NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- R01 GM086372-03/GM/NIGMS NIH HHS/United States
- R01 AI073868/AI/NIAID NIH HHS/United States
- R21 AI076921/AI/NIAID NIH HHS/United States
- R01 CA127279-04/CA/NCI NIH HHS/United States
- R01 GM086372/GM/NIGMS NIH HHS/United States
- R01 AI073868-04/AI/NIAID NIH HHS/United States
- R01 CA127279-03/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources