

Autophagy: assays and artifacts
- PMID: 20225337
- PMCID: PMC2989884
- DOI: 10.1002/path.2694
Autophagy: assays and artifacts
Abstract
Autophagy is a fundamental and phylogenetically conserved self-degradation process that is characterized by the formation of double-layered vesicles (autophagosomes) around intracellular cargo for delivery to lysosomes and proteolytic degradation. The increasing significance attached to autophagy in development and disease in higher eukaryotes has placed greater importance on the validation of reliable, meaningful and quantitative assays to monitor autophagy in live cells and in vivo in the animal. To date, the detection of processed LC3B-II by western blot or fluorescence studies, together with electron microscopy for autophagosome formation, have been the mainstays for autophagy detection. However, LC3 expression levels can vary markedly between different cell types and in response to different stresses, and there is also concern that over-expression of tagged versions of LC3 to facilitate imaging and detection of autophagy interferes with the process itself. In addition, the realization that it is not sufficient to monitor static levels of autophagy but to measure 'autophagic flux' has driven the development of new or modified approaches to detecting autophagy. Here, we present a critical overview of current methodologies to measure autophagy in cells and in animals.
Copyright (c) 2010 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Figures
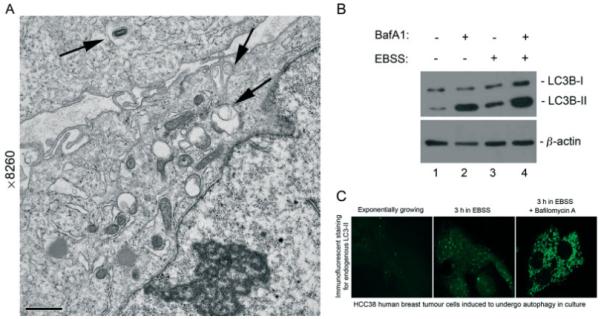
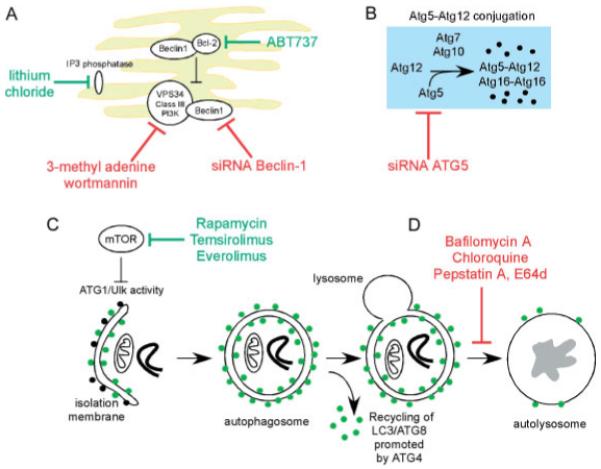
Similar articles
-
Measurement of Autophagy by Flow Cytometry.Methods Mol Biol. 2017;1553:209-216. doi: 10.1007/978-1-4939-6756-8_16. Methods Mol Biol. 2017. PMID: 28229418
-
Lysosomal turnover, but not a cellular level, of endogenous LC3 is a marker for autophagy.Autophagy. 2005 Jul;1(2):84-91. doi: 10.4161/auto.1.2.1697. Epub 2005 Jul 31. Autophagy. 2005. PMID: 16874052
-
In vivo measurement of autophagic flux by fluorescence correlation spectroscopy.Analyst. 2023 Jul 26;148(15):3498-3508. doi: 10.1039/d3an00651d. Analyst. 2023. PMID: 37409399
-
The late stages of autophagy: how does the end begin?Cell Death Differ. 2009 Jul;16(7):984-90. doi: 10.1038/cdd.2009.54. Epub 2009 May 8. Cell Death Differ. 2009. PMID: 19424283 Review.
-
Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes.Autophagy. 2008 Feb;4(2):151-75. doi: 10.4161/auto.5338. Epub 2007 Nov 21. Autophagy. 2008. PMID: 18188003 Free PMC article. Review.
Cited by
-
Involvement of macroautophagy in multiple system atrophy and protein aggregate formation in oligodendrocytes.J Mol Neurosci. 2012 Jun;47(2):256-66. doi: 10.1007/s12031-012-9733-5. Epub 2012 Mar 13. J Mol Neurosci. 2012. PMID: 22411133
-
Survival motor neuron protein reduction deregulates autophagy in spinal cord motoneurons in vitro.Cell Death Dis. 2013 Jun 20;4(6):e686. doi: 10.1038/cddis.2013.209. Cell Death Dis. 2013. PMID: 23788043 Free PMC article.
-
Superior anti-tumor efficacy of diisopropylamine dichloroacetate compared with dichloroacetate in a subcutaneous transplantation breast tumor model.Oncotarget. 2016 Oct 4;7(40):65721-65731. doi: 10.18632/oncotarget.11609. Oncotarget. 2016. PMID: 27582548 Free PMC article.
-
P300/CBP-associated factor (PCAF) inhibits the growth of hepatocellular carcinoma by promoting cell autophagy.Cell Death Dis. 2016 Oct 6;7(10):e2400. doi: 10.1038/cddis.2016.247. Cell Death Dis. 2016. PMID: 27711074 Free PMC article.
-
Human Cathelicidin Peptide LL-37 Induces Cell Death in Autophagy-Dysfunctional Endothelial Cells.J Immunol. 2022 May 1;208(9):2163-2172. doi: 10.4049/jimmunol.2100050. Epub 2022 Apr 6. J Immunol. 2022. PMID: 35387840 Free PMC article.
References
-
- Eskelinen EL. Fine structure of the autophagosome. Methods Mol Biol. 2008;445:11–28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
