

Review
doi: 10.1242/dev.038711.
To build a synapse: signaling pathways in neuromuscular junction assembly
Affiliations
- PMID: 20215342
- PMCID: PMC2835321
- DOI: 10.1242/dev.038711
Item in Clipboard
Review
To build a synapse: signaling pathways in neuromuscular junction assembly
Development.
2010 Apr.
Abstract
Synapses, as fundamental units of the neural circuitry, enable complex behaviors. The neuromuscular junction (NMJ) is a synapse type that forms between motoneurons and skeletal muscle fibers and that exhibits a high degree of subcellular specialization. Aided by genetic techniques and suitable animal models, studies in the past decade have brought significant progress in identifying NMJ components and assembly mechanisms. This review highlights recent advances in the study of NMJ development, focusing on signaling pathways that are activated by diffusible cues, which shed light on synaptogenesis in the brain and contribute to a better understanding of muscular dystrophy.
Figures
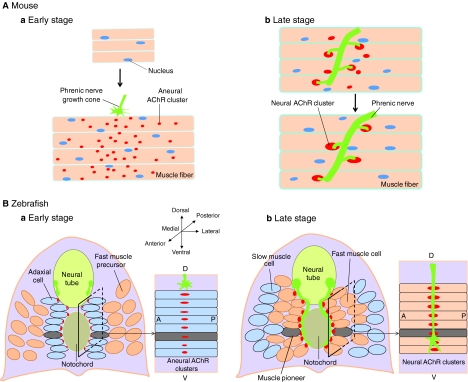
Neuromuscular synapse formation in vertebrates. (A) In mice, muscle fibers of the diaphragm form primitive, aneural AChR clusters prior to the arrival of phrenic nerve terminals. The clusters are distributed in a broad, poorly defined region in the middle of muscle fibers, a phenomenon called prepatterning (a). Innervation leads to the appearance of large AChR clusters in the synaptic region and to the disappearance of primitive clusters in non-synaptic areas (b). (B) In zebrafish, adaxial, slow muscle cells form aneural AChR clusters on the medial surface prior to innervation (a). Subsequently, adaxial muscle cells migrate outwards, and their original position is filled with fast muscle cells that form nerve-induced AChR clusters where the aneural clusters used to be (b).

Coordinated action of positive and negative signals in NMJ assembly. At least three cellular mechanisms contribute to the high density of AChRs at the NMJ. First, AChR might redistribute from primitive clusters to the synaptic area, either by lateral movement, by diffusion in the plasma membrane or by endo- and exocytosis. Second, muscle fibers are multi-nucleated cells, and only the nuclei beneath the postsynaptic membrane (synaptic nuclei) are actively transcribing the AChR subunit genes, contributing to synapse-specific AChR expression. Third, AChR turnover rate is reduced at mature NMJ or when clustered (as shown by the half-life of AChRs at the NMJ at 8-14 days compared with 17-24 hours for non-clustered or embryonic AChR). Motor nerves activate muscle fibers by releasing ACh, a negative signal, which activates AChR. Muscle activation stimulates the serine/threonine kinases cyclin-dependent kinase 5 (Cdk5) and Ca2+/calmodulin-dependent kinase II (CaMKII) to inhibit AChR clustering, to suppress AChR expression and to destabilize AChR clusters in entire muscle fibers. At the same time, nerves also release positive signals, such as agrin, which counteract the effects of negative signals, resulting in a high AChR concentration at the NMJ.

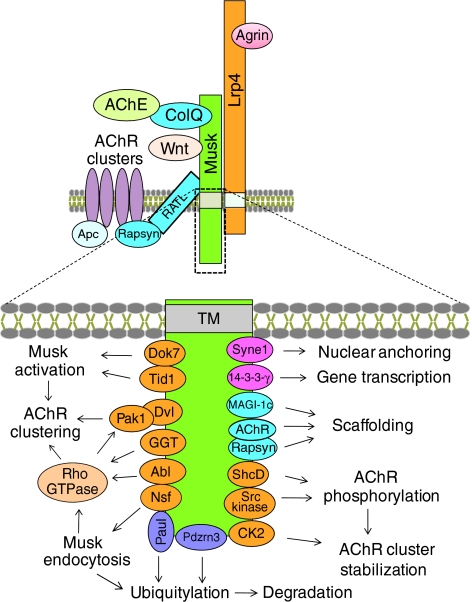
Intracellular pathways activated by agrin for AChR clustering. Agrin interacts with Lrp4 to increase its interaction with Musk and the dimerization of Musk and thus Musk activation. Subsequently, interactions between the kinase and distinct proteins, such as Dok7, which are crucial for its catalytic activity and for downstream signaling, are increased. Concomitantly, the tyrosine kinase Abl and and the metalloenzyme geranylgeranyl transferase I (GGT) are activated. GGT facilitates Rho GTPase activation, which, via multiple pathways, regulates the actin dynamics involved in AChR trafficking, membrane insertion and immobilization. Agrin also stimulates the association of AChR with rapsyn and Apc, which link the receptor directly or indirectly to the cytoskeleton. Rapsyn stability is increased at the synapse by the chaperone Hsp90β, and rapsyn interacts with and inhibits calpain, and thus antagonizes the AChR cluster-dispersing effect of ACh. Agrin signaling is regulated by Musk endocytosis, the E3 ubiquitin ligase Pdzrn3, lipid microdomains (Kishi et al., 2005; Stetzkowski-Marden et al., 2006; Willmann et al., 2006; Zhu et al., 2006) and intracellular calcium (Megeath and Fallon 1998). Many pathways illustrated here have been identified in cultured muscle and non-muscle cells, and their role in vivo remains to be studied. For example, mice lacking Shp2, a cytoplasmic tyrosine phosphatase, are viable and form normal NMJs (Dong et al., 2006), although many in vitro studies suggest a crucial role in AChR clustering. See text for details. Stars indicate protein-protein interactions that are increased by agrin. Red lipid bilayers indicate lipid rafts.

NMJ assembly in Drosophila and in C. elegans. (A) The Drosophila NMJ is a glutamatergic synapse. The Wnt ligands Wingless (Wg) and Wnt5 are released from presynaptic boutons. Binding to the Wnt receptor Frizzled on the postsynaptic membrane, Wg is thought to stimulate a Frizzled nuclear import (FNI) pathway for gene expression. This pathway might be crucial for both pre- and postsynaptic differentiation. Wg also activates Frizzled on the presynaptic membrane to promote presynaptic assembly via a pathway dependent on Gsk3β and MAP1B. Conversely, Wnt5 directly binds to and activates the receptor tyrosine kinase Derailed (Drl) on the postsynaptic membrane for postsynaptic assembly. (B) The C. elegans DA9 neuron, which is positioned in the ventral region of the worm, sends a dendrite anteriorly (green) and an axon (blue) that first migrates posteriorly; this soon turns toward the dorsal region and travels anteriorly in the dorsal region. It does not form synapses until it reaches a more anterior region, leaving an asynaptic domain in the axon. LIN-44/Wnt is produced by hypodermal cells and activates LIN-17/Frizzled in the axon to suppress presynaptic assembly.
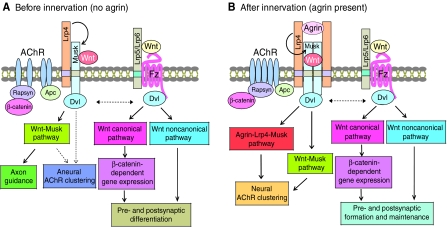
A working model of signaling pathways in postsynaptic assembly. (A) Prior to innervation, Wnt interacts with Musk to activate the Wnt-Musk pathways necessary for forming aneural clusters and for guiding motor growth cones to the middle region of muscle fibers. At the same time, Wnt might also activate Wnt canonical and non-canonical pathways to regulate Musk-dependent pathways. (B) Upon innervation, neural agrin binds to Lrp4 and activates Musk to initiate the agrin-Lrp4-Musk pathway (see Fig. 3) for AChR clustering at the synapse. Wnt might regulate agrin signaling by directly binding to Musk or Lrp5/Lrp6-Frizzled. Dashed lines indicate pathways that remain to be determined.
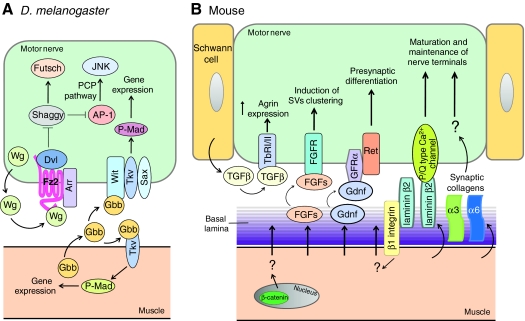
Retrograde mechanisms in NMJ formation. (A) In Drosophila, muscle cells release TGFβ/Gbb for both post- and presynaptic assembly. Wnt released from presynaptic neurons activates the Frizzled-dependent pathway for presynaptic differentiation. (B) In rodents, muscle fibers release Gdnf, which activates the receptor tyrosine kinase Ret for presynaptic differentiation. FGF, synaptic laminin and synaptic collagens orchestrate the complex temporal control of presynaptic differentiation. The interaction of laminin β2 with P/Q type calcium channels might be required for nerve terminal maturation. Muscle fibers also regulate presynaptic differentiation via a pathway that requires muscle- but not motoneuron-generated β-catenin or β1 integrin.



Similar articles
-
Glia cell line-derived neurotrophic factor regulates the distribution of acetylcholine receptors in mouse primary skeletal muscle cells.Neuroscience. 2004;128(3):497-509. doi: 10.1016/j.neuroscience.2004.06.067. Neuroscience. 2004. PMID: 15381279
-
The neuromuscular junction: selective remodeling of synaptic regulators at the nerve/muscle interface.Mech Dev. 2013 Jun-Aug;130(6-8):402-11. doi: 10.1016/j.mod.2012.09.004. Epub 2012 Sep 29. Mech Dev. 2013. PMID: 23032192 Review.
-
New dogs in the dogma: Lrp4 and Tid1 in neuromuscular synapse formation.Neuron. 2008 Nov 26;60(4):526-8. doi: 10.1016/j.neuron.2008.11.003. Neuron. 2008. PMID: 19038209 Review.
-
Molecular dissection of neuromuscular junction formation.Trends Neurosci. 2003 Jul;26(7):335-7. doi: 10.1016/S0166-2236(03)00131-0. Trends Neurosci. 2003. PMID: 12850425 Review.
-
LRP4 serves as a coreceptor of agrin.Neuron. 2008 Oct 23;60(2):285-97. doi: 10.1016/j.neuron.2008.10.006. Neuron. 2008. PMID: 18957220 Free PMC article.
Cited by
-
A role for the calmodulin kinase II-related anchoring protein (αkap) in maintaining the stability of nicotinic acetylcholine receptors.J Neurosci. 2012 Apr 11;32(15):5177-85. doi: 10.1523/JNEUROSCI.6477-11.2012. J Neurosci. 2012. PMID: 22496563 Free PMC article.
-
Slit2 as a β-catenin/Ctnnb1-dependent retrograde signal for presynaptic differentiation.Elife. 2015 Jul 10;4:e07266. doi: 10.7554/eLife.07266. Elife. 2015. PMID: 26159615 Free PMC article.
-
Assessment of the expression and role of the α1-nAChR subunit in efferent cholinergic function during the development of the mammalian cochlea.J Neurophysiol. 2016 Aug 1;116(2):479-92. doi: 10.1152/jn.01038.2015. Epub 2016 Apr 20. J Neurophysiol. 2016. PMID: 27098031 Free PMC article.
-
Serine residues in the α4 nicotinic acetylcholine receptor subunit regulate surface α4β2* receptor expression and clustering.Biochem Pharmacol. 2019 Jan;159:64-73. doi: 10.1016/j.bcp.2018.11.008. Epub 2018 Nov 9. Biochem Pharmacol. 2019. PMID: 30414940 Free PMC article.
-
Anterograde Jelly belly ligand to Alk receptor signaling at developing synapses is regulated by Mind the gap.Development. 2010 Oct;137(20):3523-33. doi: 10.1242/dev.047878. Development. 2010. PMID: 20876658 Free PMC article.
References
-
- Aberle H., Haghighi A. P., Fetter R. D., McCabe B. D., Magalhaes T. R., Goodman C. S. (2002). wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron 33, 545-558 - PubMed
-
- Akaaboune M., Culican S. M., Turney S. G., Lichtman J. W. (1999). Rapid and reversible effects of activity on acetylcholine receptor density at the neuromuscular junction in vivo. Science 286, 503-557 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
