

Structural requirements for VAP-B oligomerization and their implication in amyotrophic lateral sclerosis-associated VAP-B(P56S) neurotoxicity
- PMID: 20207736
- PMCID: PMC2859547
- DOI: 10.1074/jbc.M109.097345
Structural requirements for VAP-B oligomerization and their implication in amyotrophic lateral sclerosis-associated VAP-B(P56S) neurotoxicity
Abstract
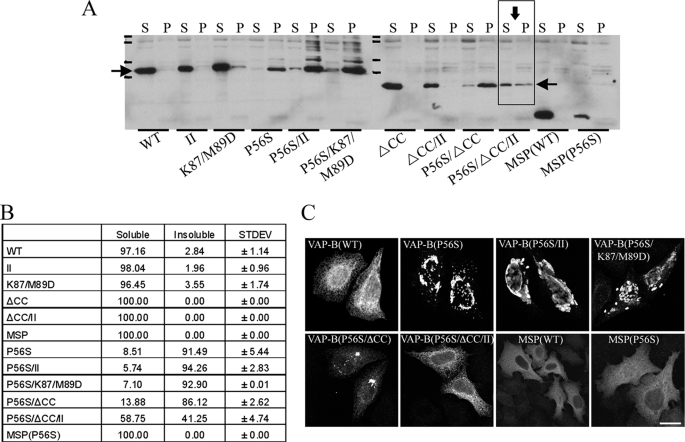
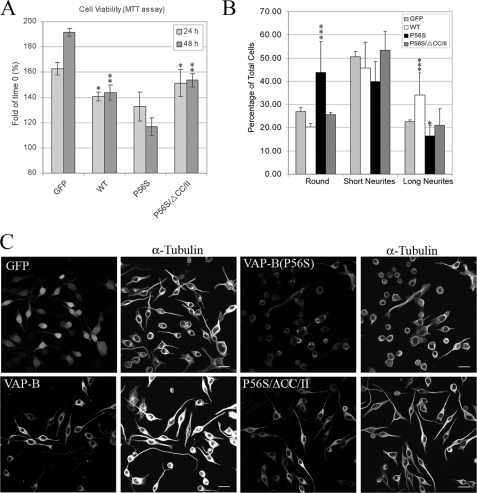
The integral endoplasmic reticulum (ER)-membrane protein VAP-B interacts with various lipid-transfer/binding proteins containing an FFAT motif through its N-terminal MSP domain. A genetic mutation within its MSP domain, P56S, was identified in familial forms of motor neuron diseases. This mutation induces the formation of insoluble VAP-B(P56S) protein aggregates by an unknown mechanism. In this study, we defined the structural requirements for VAP-B oligomerization and demonstrated their contribution for VAP-B(P56S) aggregation and neurotoxicity. We show that the oligomerization of VAP-B is mainly mediated by its coiled-coil domain and that the GXXXG dimerization motif within the transmembrane domain mediates transmembrane domains self-association but is insufficient to drive VAP-B oligomerization. We further show that the oligomerization of the wild-type VAP-B is independent of its MSP domain. However, we found that the P56S mutation induces conformational changes within the MSP domain and facilitates its propensity to aggregate by exposing hydrophobic patches to the solvent. These conformational changes have no direct effect on FFAT binding. Rather, they enhance VAP-B(P56S) oligomerization driven by the combined contributions of the coiled-coil and the transmembrane domains, thereby preventing accessibility to FFAT-binding site, facilitating the production of VAP-B(P56S)-insoluble aggregates and consequently its neurotoxicity. These results shed light on the mechanism by which VAP-B(P56S) aggregates are formed and induce familial motor neuron diseases.
Figures

Similar articles
-
A mutation in human VAP-B--MSP domain, present in ALS patients, affects the interaction with other cellular proteins.Protein Expr Purif. 2007 Sep;55(1):139-46. doi: 10.1016/j.pep.2007.04.007. Epub 2007 Apr 20. Protein Expr Purif. 2007. PMID: 17540579
-
Motor neuron disease-associated mutant vesicle-associated membrane protein-associated protein (VAP) B recruits wild-type VAPs into endoplasmic reticulum-derived tubular aggregates.J Neurosci. 2007 Sep 5;27(36):9801-15. doi: 10.1523/JNEUROSCI.2661-07.2007. J Neurosci. 2007. PMID: 17804640 Free PMC article.
-
FFAT rescues VAPA-mediated inhibition of ER-to-Golgi transport and VAPB-mediated ER aggregation.J Cell Sci. 2008 Sep 15;121(Pt 18):3052-61. doi: 10.1242/jcs.028696. Epub 2008 Aug 19. J Cell Sci. 2008. PMID: 18713837 Free PMC article.
-
The Link between VAPB Loss of Function and Amyotrophic Lateral Sclerosis.Cells. 2021 Jul 23;10(8):1865. doi: 10.3390/cells10081865. Cells. 2021. PMID: 34440634 Free PMC article. Review.
-
VAP, a Versatile Access Point for the Endoplasmic Reticulum: Review and analysis of FFAT-like motifs in the VAPome.Biochim Biophys Acta. 2016 Aug;1861(8 Pt B):952-961. doi: 10.1016/j.bbalip.2016.02.009. Epub 2016 Feb 17. Biochim Biophys Acta. 2016. PMID: 26898182 Review.
Cited by
-
VAPB ER-Aggregates, A Possible New Biomarker in ALS Pathology.Cells. 2020 Jan 9;9(1):164. doi: 10.3390/cells9010164. Cells. 2020. PMID: 31936602 Free PMC article.
-
Downregulation of VAPB expression in motor neurons derived from induced pluripotent stem cells of ALS8 patients.Hum Mol Genet. 2011 Sep 15;20(18):3642-52. doi: 10.1093/hmg/ddr284. Epub 2011 Jun 17. Hum Mol Genet. 2011. PMID: 21685205 Free PMC article.
-
ER-PM Junctions on GABAergic Interneurons Are Organized by Neuregulin 2/VAP Interactions and Regulated by NMDA Receptors.Int J Mol Sci. 2023 Feb 2;24(3):2908. doi: 10.3390/ijms24032908. Int J Mol Sci. 2023. PMID: 36769244 Free PMC article.
-
Amyotrophic lateral sclerosis-associated mutant VAPBP56S perturbs calcium homeostasis to disrupt axonal transport of mitochondria.Hum Mol Genet. 2012 May 1;21(9):1979-88. doi: 10.1093/hmg/dds011. Epub 2012 Jan 17. Hum Mol Genet. 2012. PMID: 22258555 Free PMC article.
-
Analysis of the key elements of FFAT-like motifs identifies new proteins that potentially bind VAP on the ER, including two AKAPs and FAPP2.PLoS One. 2012;7(1):e30455. doi: 10.1371/journal.pone.0030455. Epub 2012 Jan 19. PLoS One. 2012. PMID: 22276202 Free PMC article.
References
-
- Ross C. A., Poirier M. A. (2004) Nat. Med. 10, (suppl.) S10–S17 - PubMed
-
- Amarilio R., Ramachandran S., Sabanay H., Lev S. (2005) J. Biol. Chem. 280, 5934–5944 - PubMed
-
- Brown R. H., Jr., Robberecht W. (2001) Semin. Neurol. 21, 131–139 - PubMed
-
- Orrell R. W. (2000) Neuromuscul. Disord. 10, 63–68 - PubMed
-
- Gros-Louis F., Gaspar C., Rouleau G. A. (2006) Biochim. Biophys. Acta 1762, 956–972 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
