

Orm family proteins mediate sphingolipid homeostasis
- PMID: 20182505
- PMCID: PMC2877384
- DOI: 10.1038/nature08787
Orm family proteins mediate sphingolipid homeostasis
Abstract
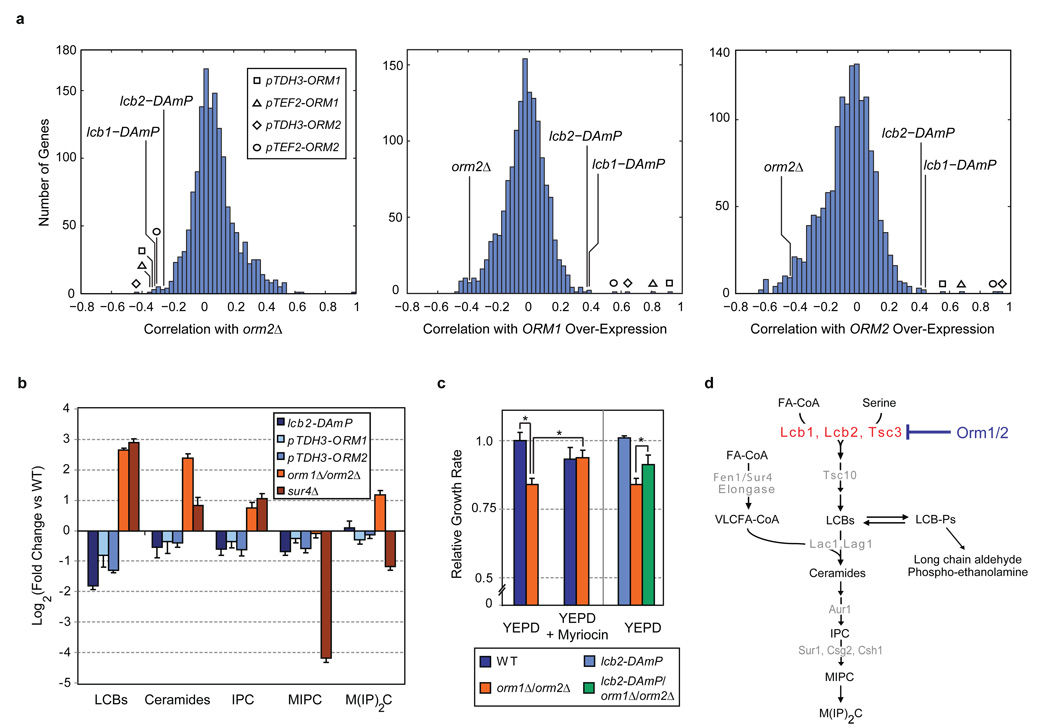
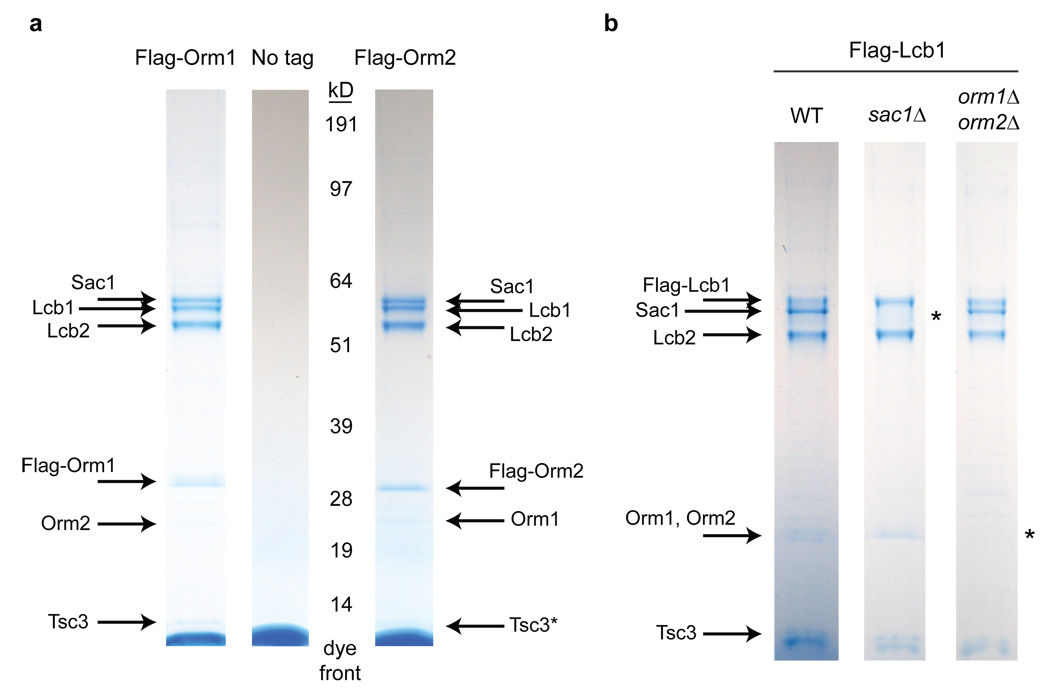
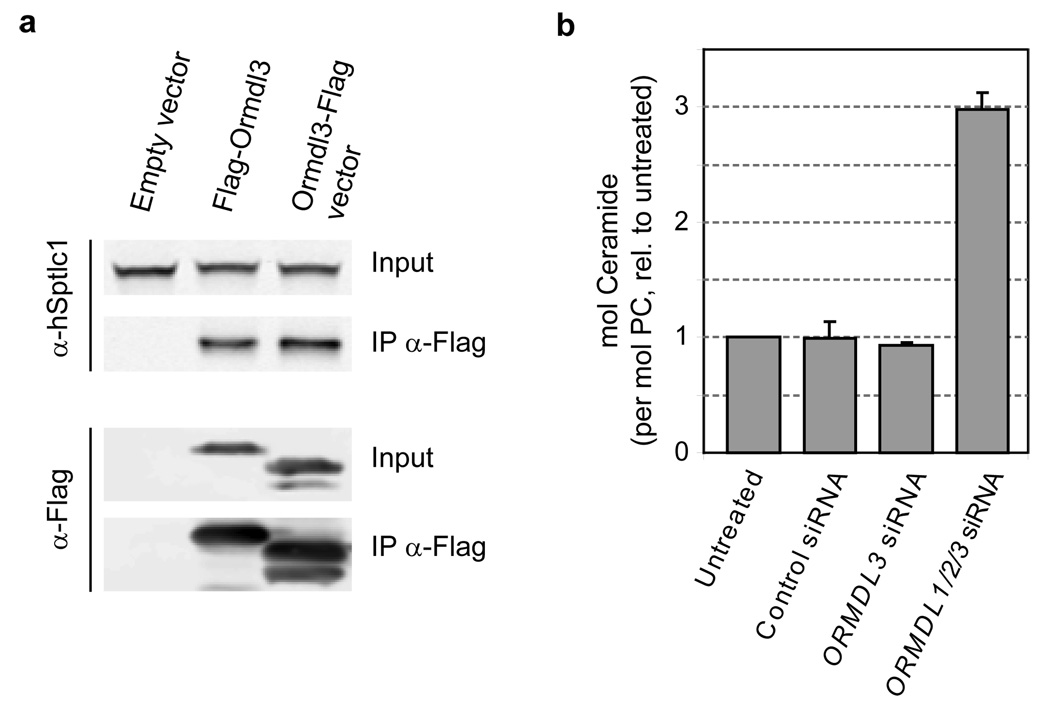
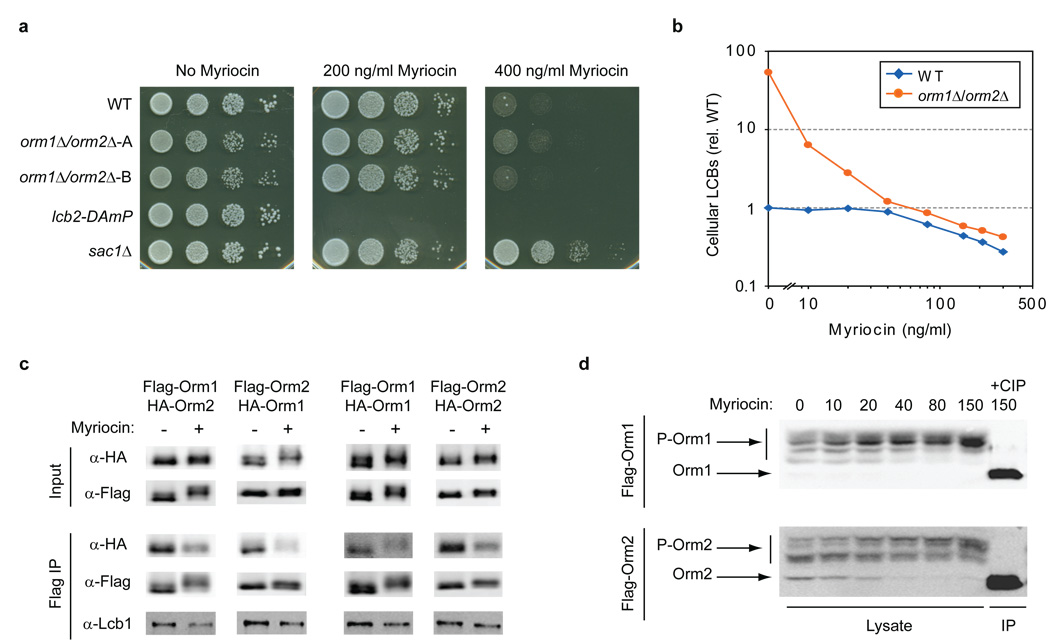
Despite the essential roles of sphingolipids both as structural components of membranes and critical signalling molecules, we have a limited understanding of how cells sense and regulate their levels. Here we reveal the function in sphingolipid metabolism of the ORM genes (known as ORMDL genes in humans)-a conserved gene family that includes ORMDL3, which has recently been identified as a potential risk factor for childhood asthma. Starting from an unbiased functional genomic approach in Saccharomyces cerevisiae, we identify Orm proteins as negative regulators of sphingolipid synthesis that form a conserved complex with serine palmitoyltransferase, the first and rate-limiting enzyme in sphingolipid production. We also define a regulatory pathway in which phosphorylation of Orm proteins relieves their inhibitory activity when sphingolipid production is disrupted. Changes in ORM gene expression or mutations to their phosphorylation sites cause dysregulation of sphingolipid metabolism. Our work identifies the Orm proteins as critical mediators of sphingolipid homeostasis and raises the possibility that sphingolipid misregulation contributes to the development of childhood asthma.
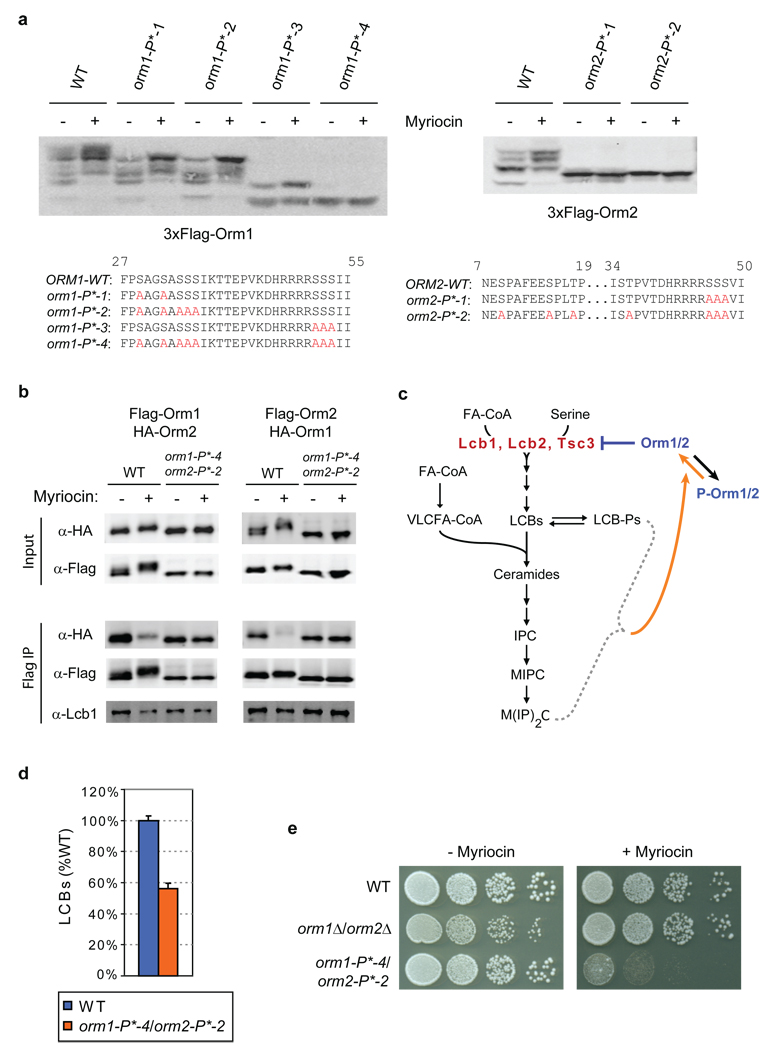
Figures
Comment in
-
Cell biology: A brake on lipid synthesis.Nature. 2010 Feb 25;463(7284):1028-9. doi: 10.1038/4631028a. Nature. 2010. PMID: 20182499 No abstract available.
Similar articles
-
Orm proteins integrate multiple signals to maintain sphingolipid homeostasis.J Biol Chem. 2013 Jul 12;288(28):20453-63. doi: 10.1074/jbc.M113.472860. Epub 2013 Jun 4. J Biol Chem. 2013. PMID: 23737533 Free PMC article.
-
Protein kinase Ypk1 phosphorylates regulatory proteins Orm1 and Orm2 to control sphingolipid homeostasis in Saccharomyces cerevisiae.Proc Natl Acad Sci U S A. 2011 Nov 29;108(48):19222-7. doi: 10.1073/pnas.1116948108. Epub 2011 Nov 11. Proc Natl Acad Sci U S A. 2011. PMID: 22080611 Free PMC article.
-
TORC1-regulated protein kinase Npr1 phosphorylates Orm to stimulate complex sphingolipid synthesis.Mol Biol Cell. 2013 Mar;24(6):870-81. doi: 10.1091/mbc.E12-10-0753. Epub 2013 Jan 30. Mol Biol Cell. 2013. PMID: 23363605 Free PMC article.
-
Orm/ORMDL proteins: Gate guardians and master regulators.Adv Biol Regul. 2018 Dec;70:3-18. doi: 10.1016/j.jbior.2018.08.002. Epub 2018 Aug 31. Adv Biol Regul. 2018. PMID: 30193828 Free PMC article. Review.
-
The role of ORMDL proteins, guardians of cellular sphingolipids, in asthma.Allergy. 2016 Jul;71(7):918-30. doi: 10.1111/all.12877. Epub 2016 Mar 29. Allergy. 2016. PMID: 26969910 Review.
Cited by
-
TOR complex 2 is a master regulator of plasma membrane homeostasis.Biochem J. 2022 Sep 30;479(18):1917-1940. doi: 10.1042/BCJ20220388. Biochem J. 2022. PMID: 36149412 Free PMC article.
-
Flavonoids as Potential Drugs for VPS13-Dependent Rare Neurodegenerative Diseases.Genes (Basel). 2020 Jul 21;11(7):828. doi: 10.3390/genes11070828. Genes (Basel). 2020. PMID: 32708255 Free PMC article.
-
Tubular ER Associates With Diacylglycerol-Rich Structures During Lipid Droplet Consumption.Front Cell Dev Biol. 2020 Jul 29;8:700. doi: 10.3389/fcell.2020.00700. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32850820 Free PMC article.
-
Dietary long-chain omega 3 fatty acids modify sphingolipid metabolism to facilitate airway hyperreactivity.Sci Rep. 2022 Nov 17;12(1):19735. doi: 10.1038/s41598-022-21083-w. Sci Rep. 2022. PMID: 36396956 Free PMC article.
-
Palmitate and lipopolysaccharide trigger synergistic ceramide production in primary macrophages.J Biol Chem. 2013 Feb 1;288(5):2923-32. doi: 10.1074/jbc.M112.419978. Epub 2012 Dec 18. J Biol Chem. 2013. PMID: 23250746 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
