

LSm1-7 complexes bind to specific sites in viral RNA genomes and regulate their translation and replication
- PMID: 20181739
- PMCID: PMC2844628
- DOI: 10.1261/rna.1712910
LSm1-7 complexes bind to specific sites in viral RNA genomes and regulate their translation and replication
Abstract
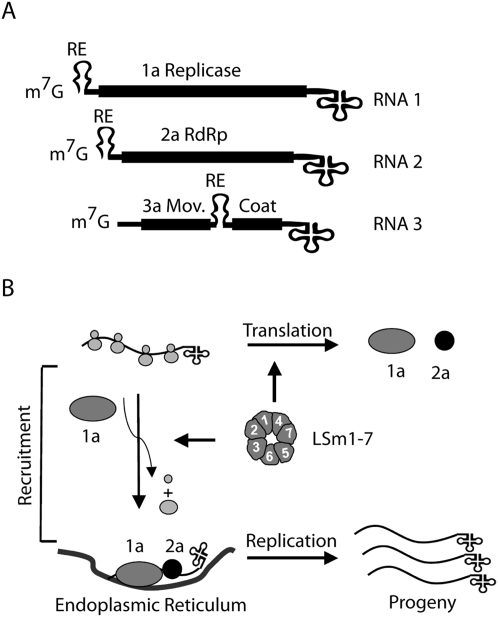
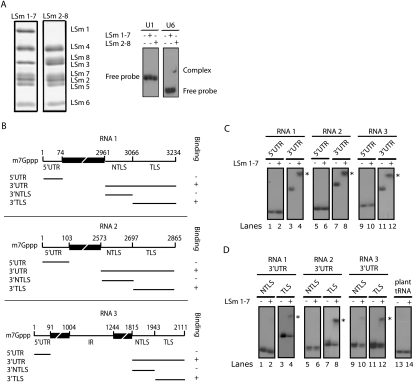
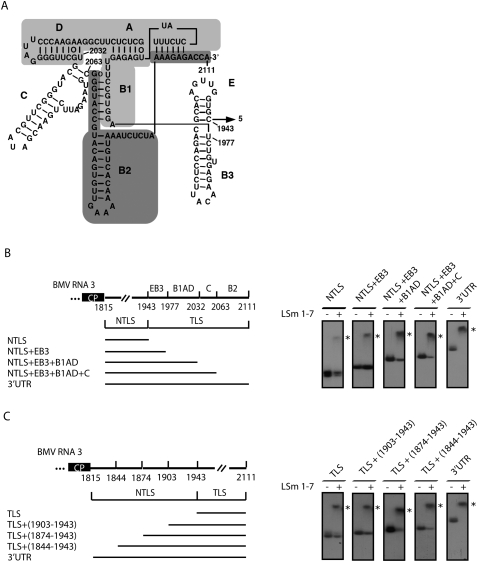
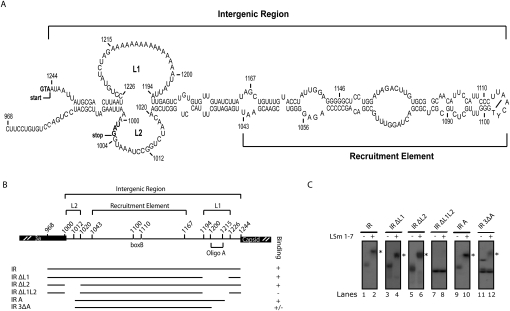
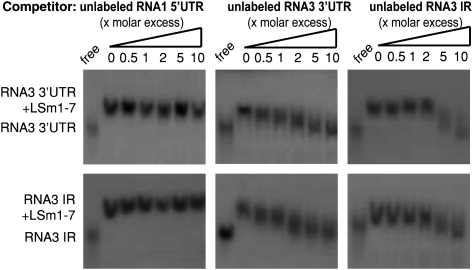
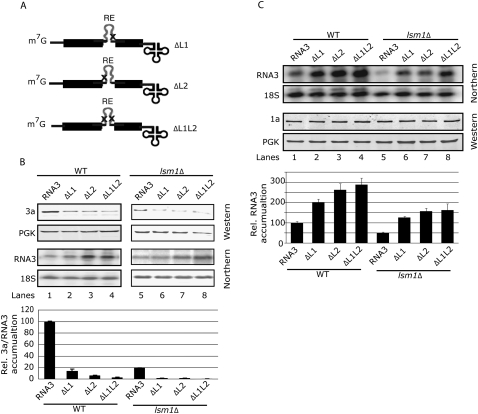
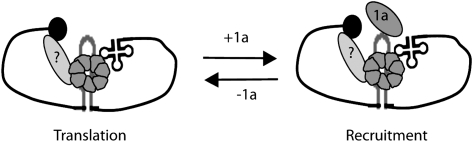
LSm1-7 complexes promote cellular mRNA degradation, in addition to translation and replication of positive-strand RNA viruses such as the Brome mosaic virus (BMV). Yet, how LSm1-7 complexes act on their targets remains elusive. Here, we report that reconstituted recombinant LSm1-7 complexes directly bind to two distinct RNA-target sequences in the BMV genome, a tRNA-like structure at the 3'-untranslated region and two internal A-rich single-stranded regions. Importantly, in vivo analysis shows that these sequences regulate the translation and replication of the BMV genome. Furthermore, both RNA-target sequences resemble those found for Hfq, the LSm counterpart in bacteria, suggesting conservation through evolution. Our results provide the first evidence that LSm1-7 complexes interact directly with viral RNA genomes and open new perspectives in the understanding of LSm1-7 functions.
Figures

Similar articles
-
The Lsm1-7-Pat1 complex promotes viral RNA translation and replication by differential mechanisms.RNA. 2015 Aug;21(8):1469-79. doi: 10.1261/rna.052209.115. Epub 2015 Jun 19. RNA. 2015. PMID: 26092942 Free PMC article.
-
Yeast Lsm1p-7p/Pat1p deadenylation-dependent mRNA-decapping factors are required for brome mosaic virus genomic RNA translation.Mol Cell Biol. 2003 Jun;23(12):4094-106. doi: 10.1128/MCB.23.12.4094-4106.2003. Mol Cell Biol. 2003. PMID: 12773554 Free PMC article.
-
The cellular decapping activators LSm1, Pat1, and Dhh1 control the ratio of subgenomic to genomic Flock House virus RNAs.J Virol. 2013 Jun;87(11):6192-200. doi: 10.1128/JVI.03327-12. Epub 2013 Mar 27. J Virol. 2013. PMID: 23536653 Free PMC article.
-
The brome mosaic virus 3' untranslated sequence regulates RNA replication, recombination, and virion assembly.Virus Res. 2015 Aug 3;206:46-52. doi: 10.1016/j.virusres.2015.02.007. Epub 2015 Feb 14. Virus Res. 2015. PMID: 25687214 Review.
-
The coat protein leads the way: an update on basic and applied studies with the Brome mosaic virus coat protein.Mol Plant Pathol. 2011 May;12(4):403-12. doi: 10.1111/j.1364-3703.2010.00678.x. Epub 2010 Nov 25. Mol Plant Pathol. 2011. PMID: 21453435 Free PMC article. Review.
Cited by
-
Strategies for viral RNA stability: live long and prosper.Trends Genet. 2011 Jul;27(7):286-93. doi: 10.1016/j.tig.2011.04.003. Trends Genet. 2011. PMID: 21640425 Free PMC article. Review.
-
Plant RNA Regulatory Network and RNA Granules in Virus Infection.Front Plant Sci. 2017 Dec 11;8:2093. doi: 10.3389/fpls.2017.02093. eCollection 2017. Front Plant Sci. 2017. PMID: 29312371 Free PMC article. Review.
-
DEAD-box helicases modulate dicing body formation in Arabidopsis.Sci Adv. 2021 Apr 28;7(18):eabc6266. doi: 10.1126/sciadv.abc6266. Print 2021 Apr. Sci Adv. 2021. PMID: 33910901 Free PMC article.
-
The Lsm1-7-Pat1 complex promotes viral RNA translation and replication by differential mechanisms.RNA. 2015 Aug;21(8):1469-79. doi: 10.1261/rna.052209.115. Epub 2015 Jun 19. RNA. 2015. PMID: 26092942 Free PMC article.
-
Biomolecular condensates in plant RNA silencing: insights into formation, function, and stress responses.Plant Cell. 2024 Jan 30;36(2):227-245. doi: 10.1093/plcell/koad254. Plant Cell. 2024. PMID: 37772963 Free PMC article. Review.
References
-
- Ahlquist P. Bromovirus RNA replication and transcription. Curr Opin Genet Dev. 1992;2:71–76. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases