

A novel glucosyltransferase is required for glycosylation of a serine-rich adhesin and biofilm formation by Streptococcus parasanguinis
- PMID: 20164186
- PMCID: PMC2852953
- DOI: 10.1074/jbc.M109.066928
A novel glucosyltransferase is required for glycosylation of a serine-rich adhesin and biofilm formation by Streptococcus parasanguinis
Abstract
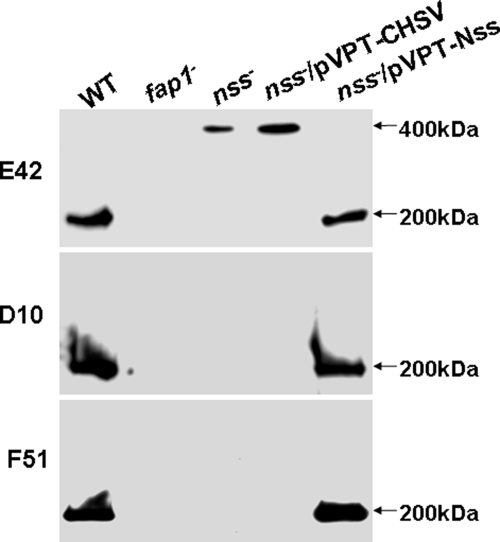
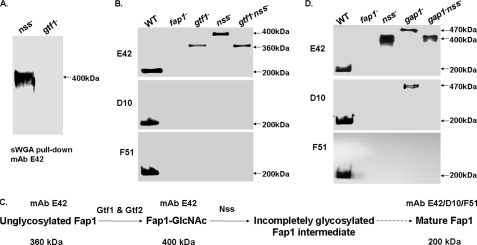
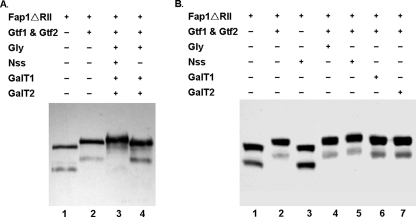
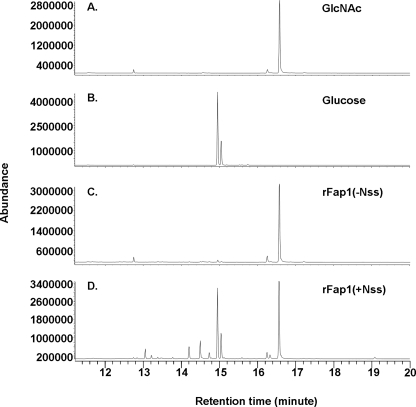
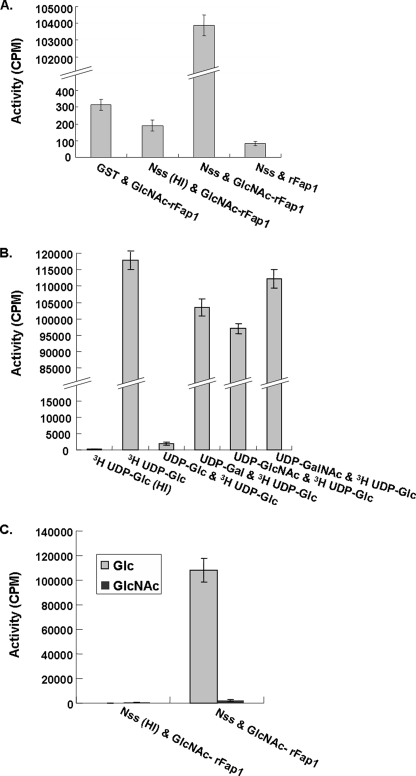
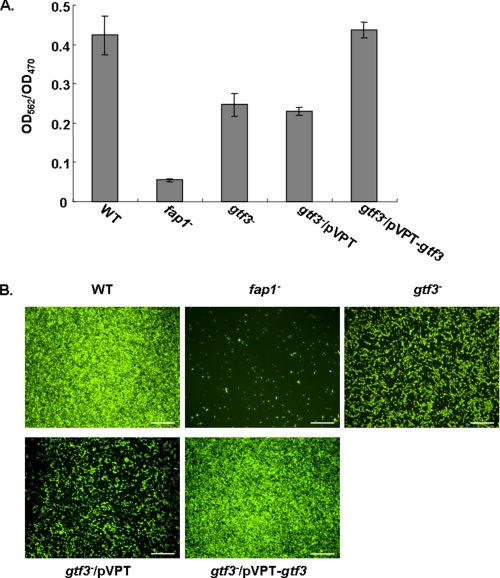
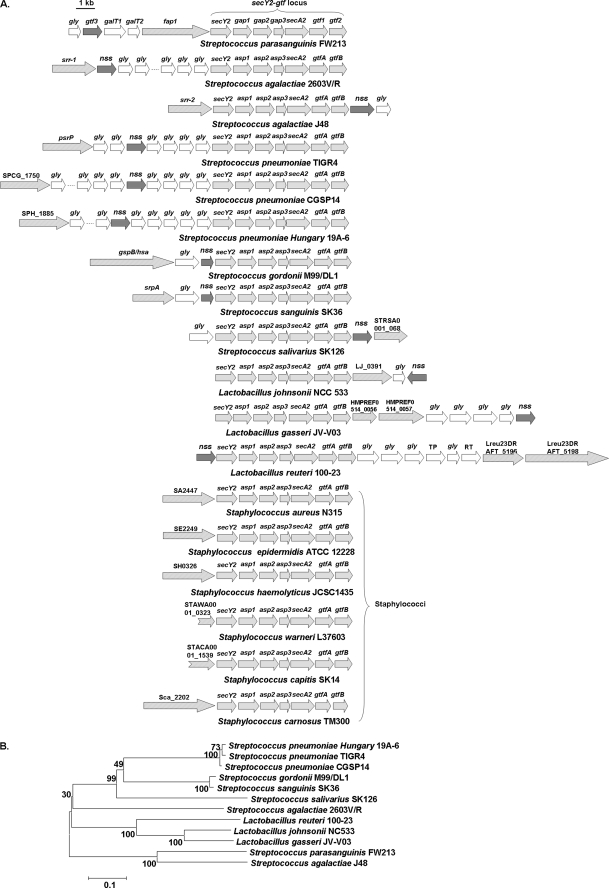
Fap1-like serine-rich glycoproteins are conserved in streptococci, staphylococci, and lactobacilli, and are required for bacterial biofilm formation and pathogenesis. Glycosylation of Fap1 is mediated by a gene cluster flanking the fap1 locus. The key enzymes responsible for the first step of Fap1 glycosylation are glycosyltransferases Gtf1 and Gtf2. They form a functional enzyme complex that catalyzes the transfer of N-acetylglucosamine (GlcNAc) residues to the Fap1 polypeptide. However, until now nothing was known about the subsequent step in Fap1 glycosylation. Here, we show that the second step in Fap1 glycosylation is catalyzed by nucleotide-sugar synthetase-like (Nss) protein. The nss gene located upstream of fap1 is also highly conserved in streptococci and lactobacilli. Nss-deficient mutants failed to catalyze the second step of Fap1 glycosylation in vivo in Streptococcus parasanguinis and in a recombinant Fap1 glycosylation system. Nss catalyzed the direct transfer of the glucosyl residue to the GlcNAc-modified Fap1 substrate in vitro, demonstrating that Nss is a glucosyltransferase. Thus we renamed Nss as glucosyltransferase 3 (Gtf3). A gtf3 mutant exhibited a biofilm defect. Taken together, we conclude that this new glucosyltransferase mediates the second step of Fap1 glycosylation and is required for biofilm formation.
Figures
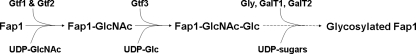
Similar articles
-
Glycosyltransferase-mediated Sweet Modification in Oral Streptococci.J Dent Res. 2015 May;94(5):659-65. doi: 10.1177/0022034515574865. Epub 2015 Mar 9. J Dent Res. 2015. PMID: 25755271 Free PMC article. Review.
-
Interaction between two putative glycosyltransferases is required for glycosylation of a serine-rich streptococcal adhesin.J Bacteriol. 2008 Feb;190(4):1256-66. doi: 10.1128/JB.01078-07. Epub 2007 Dec 14. J Bacteriol. 2008. PMID: 18083807 Free PMC article.
-
Structural and functional analysis of a new subfamily of glycosyltransferases required for glycosylation of serine-rich streptococcal adhesins.J Biol Chem. 2011 Jul 29;286(30):27048-57. doi: 10.1074/jbc.M110.208629. Epub 2011 Jun 7. J Biol Chem. 2011. PMID: 21653318 Free PMC article.
-
Engineering and Dissecting the Glycosylation Pathway of a Streptococcal Serine-rich Repeat Adhesin.J Biol Chem. 2016 Dec 30;291(53):27354-27363. doi: 10.1074/jbc.M116.752998. J Biol Chem. 2016. PMID: 28039332 Free PMC article.
-
Glycosylation and biogenesis of a family of serine-rich bacterial adhesins.Microbiology (Reading). 2009 Feb;155(Pt 2):317-327. doi: 10.1099/mic.0.025221-0. Microbiology (Reading). 2009. PMID: 19202081 Review.
Cited by
-
Glycosyltransferase-mediated Sweet Modification in Oral Streptococci.J Dent Res. 2015 May;94(5):659-65. doi: 10.1177/0022034515574865. Epub 2015 Mar 9. J Dent Res. 2015. PMID: 25755271 Free PMC article. Review.
-
A role for glycosylated serine-rich repeat proteins in gram-positive bacterial pathogenesis.Mol Oral Microbiol. 2012 Aug;27(4):257-69. doi: 10.1111/j.2041-1014.2012.00653.x. Epub 2012 Jun 11. Mol Oral Microbiol. 2012. PMID: 22759311 Free PMC article. Review.
-
Pneumococcal Surface Proteins as Virulence Factors, Immunogens, and Conserved Vaccine Targets.Front Cell Infect Microbiol. 2022 May 12;12:832254. doi: 10.3389/fcimb.2022.832254. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35646747 Free PMC article. Review.
-
Recognition of specific sialoglycan structures by oral streptococci impacts the severity of endocardial infection.PLoS Pathog. 2019 Jun 24;15(6):e1007896. doi: 10.1371/journal.ppat.1007896. eCollection 2019 Jun. PLoS Pathog. 2019. PMID: 31233555 Free PMC article.
-
The highly conserved domain of unknown function 1792 has a distinct glycosyltransferase fold.Nat Commun. 2014 Jul 15;5:4339. doi: 10.1038/ncomms5339. Nat Commun. 2014. PMID: 25023666 Free PMC article.
References
-
- Drickamer K., Taylor M. E. (1998) Trends Biochem. Sci. 23, 321–324 - PubMed
-
- Ohtsubo K., Marth J. D. (2006) Cell 126, 855–867 - PubMed
-
- Varki A. (2006) Cell 126, 841–845 - PubMed
-
- Moens S., Vanderleyden J. (1997) Arch. Microbiol. 168, 169–175 - PubMed
-
- Schmidt M. A., Riley L. W., Benz I. (2003) Trends Microbiol. 11, 554–561 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
