

doi: 10.1073/pnas.1000398107.
Epub 2010 Feb 8.
Mechanism of chromatin remodeling
Affiliations
- PMID: 20142505
- PMCID: PMC2817641
- DOI: 10.1073/pnas.1000398107
Item in Clipboard
Mechanism of chromatin remodeling
Proc Natl Acad Sci U S A.
.
Abstract
Results from biochemical and structural studies of the RSC chromatin-remodeling complex prompt a proposal for the remodeling mechanism: RSC binding to the nucleosome releases the DNA from the histone surface and initiates DNA translocation (through one or a small number of DNA base pairs); ATP binding completes translocation, and ATP hydrolysis resets the system. Binding energy thus plays a central role in the remodeling process. RSC may disrupt histone-DNA contacts by affecting histone octamer conformation and through extensive interaction with the DNA. Bulging of the DNA from the octamer surface is possible, and twisting is unavoidable, but neither is the basis of remodeling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Effects of RSC- and ATPγS binding upon DNase I digestion of nucleosomal DNA. (A) DNase I digestion patterns of naked DNA and nucleosomes, under the conditions indicated above the lanes. Digestion was with 0.33 units of DNase I (ProMega) for 15 sec (lanes 1 and 2) or with 0.6 units for 40 sec (lanes 3–8). Positions of size markers produced by digestion of naked DNA with either Msp I or Alu I are indicated by DNA length on the right. (B) Profile of radioactivity in lane 4 of the region indicated by the bracket on the right of (A). Arrows identify cutting sites exposed by RSC–nucleosome interaction. (C) DNase I digestion pattern in the vicinity of the hypersensitive site indicated by the arrow in (A), under the conditions indicated above the lanes. In this case, the nucleosomal DNA was labeled at the end opposite to that in (A). Amounts of enzyme and times of digestion were the same as in (A).

Effect of RSC-binding upon exo III digestion of nucleosomal DNA. Digestion of nucleosomes (2.5 ng DNA) was performed with 40 u exo III (New England Biolabs) for 5 min under the conditions indicated above the lanes (0.3 μg RSC, 0.5 mM ATP or ATPγS). Position of size marker produced by digestion of naked DNA with Msp I is indicated by DNA length on the right. Product of limit exo III digestion, approximately half the length of the starting nucleosomal DNA, is indicated by an asterisk.
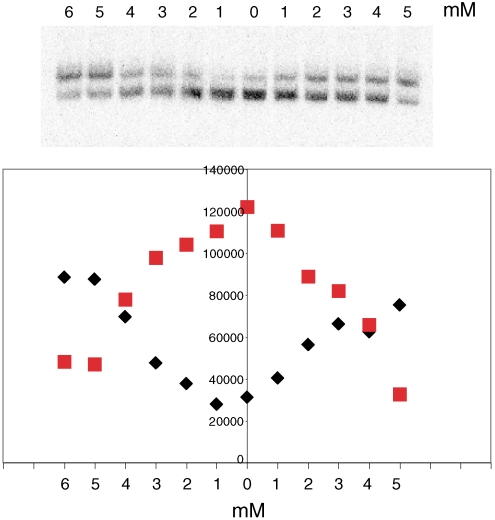
Dissociation of H2A-H2B dimers from nucleosomes and reversal in the presence of acetate. Nucleosomes (1 ng DNA) were incubated with naked DNA (1 μg pUC19) for 5 min and analyzed by gel electrophoresis. For lanes to the left of the center line, incubation was in the presence of potassium acetate at the concentrations indicated; for lanes to the right of the center line, incubation was with naked DNA alone, followed by the addition of potassium acetate at the concentrations indicated. The bands in the gel correspond to the nucleosome and tetramer particle. The relative intensities are plotted as black diamonds and red squares below.
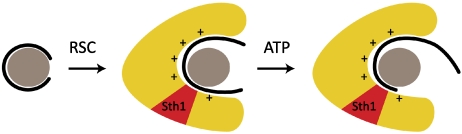
Proposed mechanism of chromatin remodeling. Schematic of nucleosome at left depicts histone octamer as gray disk and DNA as solid curve. Schematic of RSC–nucleosome complex in center depicts RSC as yellow volume surrounding the nucleosome, with Sth1 region in red, with positively charged surface for interaction with the nucleosome, and with DNA largely dissociated from surface of the histone octamer. Schematic of RSC–nucleosome at right shows DNA translocated in the presence of ATP.
Similar articles
-
Distortion of histone octamer core promotes nucleosome mobilization by a chromatin remodeler.Science. 2017 Jan 20;355(6322):eaaa3761. doi: 10.1126/science.aaa3761. Science. 2017. PMID: 28104838 Free PMC article.
-
The synergy between RSC, Nap1 and adjacent nucleosome in nucleosome remodeling.Biochim Biophys Acta Gene Regul Mech. 2019 Feb;1862(2):129-140. doi: 10.1016/j.bbagrm.2018.11.008. Epub 2018 Dec 26. Biochim Biophys Acta Gene Regul Mech. 2019. PMID: 30593928
-
Chromatin remodeling by RSC involves ATP-dependent DNA translocation.Genes Dev. 2002 Aug 15;16(16):2120-34. doi: 10.1101/gad.995002. Genes Dev. 2002. PMID: 12183366 Free PMC article.
-
ATP-dependent chromatin remodeling and DNA double-strand break repair.Cell Cycle. 2005 Aug;4(8):1011-4. doi: 10.4161/cc.4.8.1887. Epub 2005 Aug 2. Cell Cycle. 2005. PMID: 16082209 Review.
-
Snf2-family proteins: chromatin remodellers for any occasion.Curr Opin Chem Biol. 2011 Oct;15(5):649-56. doi: 10.1016/j.cbpa.2011.07.022. Epub 2011 Aug 20. Curr Opin Chem Biol. 2011. PMID: 21862382 Free PMC article. Review.
Cited by
-
Imaging of DNA and Protein-DNA Complexes with Atomic Force Microscopy.Crit Rev Eukaryot Gene Expr. 2016;26(1):63-96. doi: 10.1615/CritRevEukaryotGeneExpr.v26.i1.70. Crit Rev Eukaryot Gene Expr. 2016. PMID: 27278886 Free PMC article. Review.
-
Pyruvate Kinase M2 serves as blockade for nucleosome repositioning and abrogates Chd7 remodeling activity.PLoS One. 2019 Feb 8;14(2):e0211515. doi: 10.1371/journal.pone.0211515. eCollection 2019. PLoS One. 2019. PMID: 30735509 Free PMC article.
-
Asymmetric nucleosomes flank promoters in the budding yeast genome.Genome Res. 2015 Mar;25(3):381-90. doi: 10.1101/gr.182618.114. Epub 2014 Dec 9. Genome Res. 2015. PMID: 25491770 Free PMC article.
-
Chromatin remodelers: We are the drivers!!Nucleus. 2016 Jul 3;7(4):388-404. doi: 10.1080/19491034.2016.1211217. Epub 2016 Jul 18. Nucleus. 2016. PMID: 27429206 Free PMC article. Review.
-
Charge state of the globular histone core controls stability of the nucleosome.Biophys J. 2010 Sep 8;99(5):1577-85. doi: 10.1016/j.bpj.2010.06.046. Biophys J. 2010. PMID: 20816070 Free PMC article.
References
-
- Lorch Y, Zhang M, Kornberg RD. Histone octamer transfer by a chromatin-remodeling complex. Cell. 1999;96(3):389–392. - PubMed
-
- Whitehouse I, et al. Nucleosome mobilization catalysed by the yeast SWI/SNF complex. Nature. 1999;400(6746):784–787. - PubMed
-
- Saha A, Wittmeyer J, Cairns BR. Chromatin remodelling: The industrial revolution of DNA around histones. Nat Rev Mol Cell Biol. 2006;7(6):437–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
