

Mutations in Grxcr1 are the basis for inner ear dysfunction in the pirouette mouse
- PMID: 20137774
- PMCID: PMC2820167
- DOI: 10.1016/j.ajhg.2010.01.016
Mutations in Grxcr1 are the basis for inner ear dysfunction in the pirouette mouse
Abstract
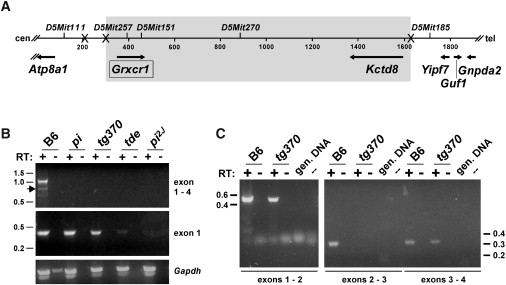
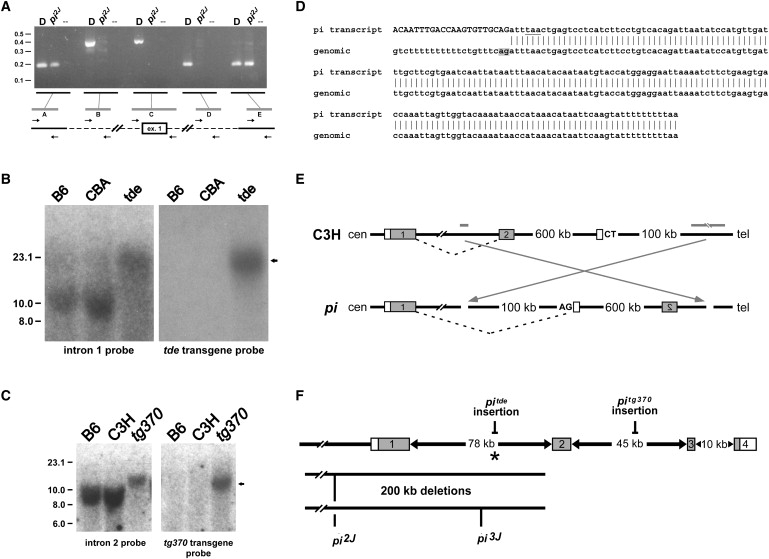
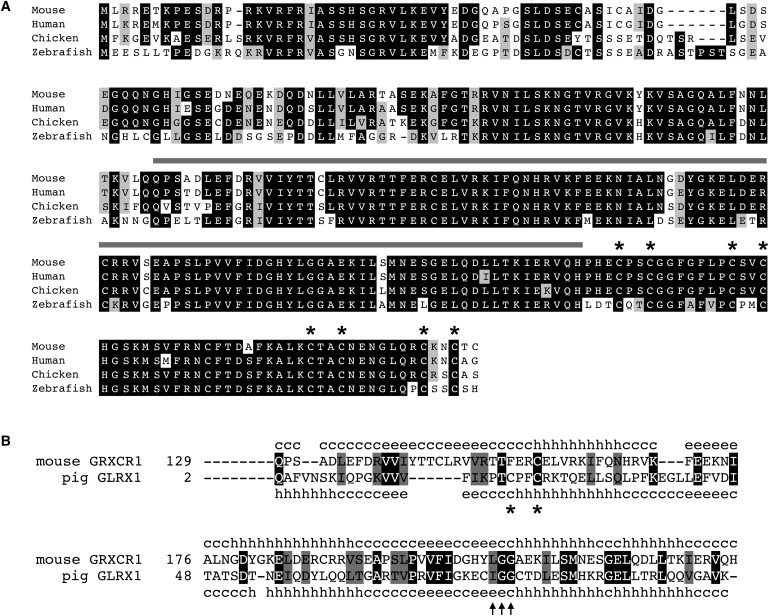
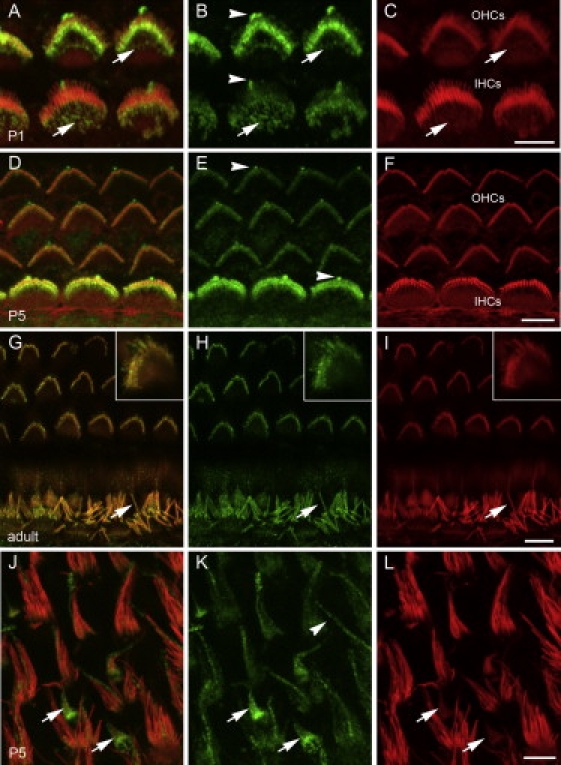
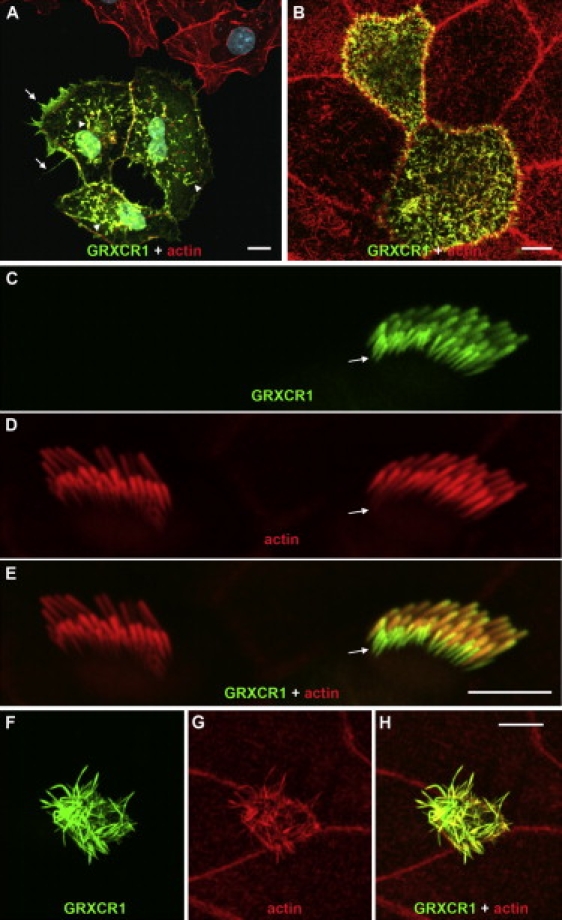
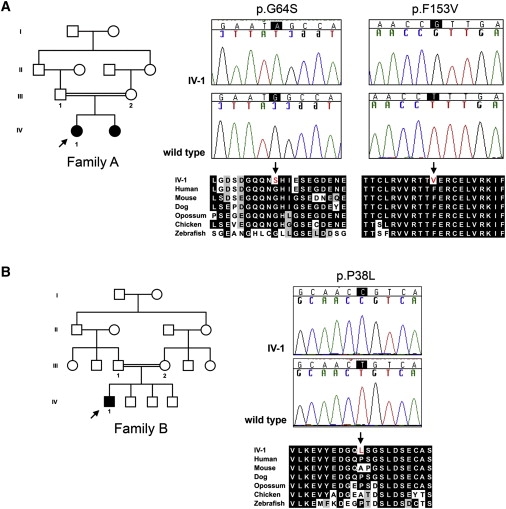
Recessive mutations at the mouse pirouette (pi) locus result in hearing loss and vestibular dysfunction due to neuroepithelial defects in the inner ear. Using a positional cloning strategy, we have identified mutations in the gene Grxcr1 (glutaredoxin cysteine-rich 1) in five independent allelic strains of pirouette mice. We also provide sequence data of GRXCR1 from humans with profound hearing loss suggesting that pirouette is a model for studying the mechanism of nonsyndromic deafness DFNB25. Grxcr1 encodes a 290 amino acid protein that contains a region of similarity to glutaredoxin proteins and a cysteine-rich region at its C terminus. Grxcr1 is expressed in sensory epithelia of the inner ear, and its encoded protein is localized along the length of stereocilia, the actin-filament-rich mechanosensory structures at the apical surface of auditory and vestibular hair cells. The precise architecture of hair cell stereocilia is essential for normal hearing. Loss of function of Grxcr1 in homozygous pirouette mice results in abnormally thin and slightly shortened stereocilia. When overexpressed in transfected cells, GRXCR1 localizes along the length of actin-filament-rich structures at the dorsal-apical surface and induces structures with greater actin filament content and/or increased lengths in a subset of cells. Our results suggest that deafness in pirouette mutants is associated with loss of GRXCR1 function in modulating actin cytoskeletal architecture in the developing stereocilia of sensory hair cells.
Copyright (c) 2010 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Homozygosity mapping reveals mutations of GRXCR1 as a cause of autosomal-recessive nonsyndromic hearing impairment.Am J Hum Genet. 2010 Feb 12;86(2):138-47. doi: 10.1016/j.ajhg.2009.12.017. Epub 2010 Feb 4. Am J Hum Genet. 2010. PMID: 20137778 Free PMC article.
-
Grxcr2 is required for stereocilia morphogenesis in the cochlea.PLoS One. 2018 Aug 29;13(8):e0201713. doi: 10.1371/journal.pone.0201713. eCollection 2018. PLoS One. 2018. PMID: 30157177 Free PMC article.
-
Characterization of two transgene insertional mutations at pirouette, a mouse deafness locus.Audiol Neurootol. 2004 Sep-Oct;9(5):303-14. doi: 10.1159/000080701. Epub 2004 Sep 3. Audiol Neurootol. 2004. PMID: 15347914
-
MicroRNAs and epigenetic regulation in the mammalian inner ear: implications for deafness.Mamm Genome. 2009 Sep-Oct;20(9-10):581-603. doi: 10.1007/s00335-009-9230-5. Epub 2009 Oct 30. Mamm Genome. 2009. PMID: 19876605 Review.
-
Gap junctions and connexins in the inner ear: their roles in homeostasis and deafness.Curr Opin Otolaryngol Head Neck Surg. 2008 Oct;16(5):452-7. doi: 10.1097/MOO.0b013e32830e20b0. Curr Opin Otolaryngol Head Neck Surg. 2008. PMID: 18797288 Review.
Cited by
-
Integrating the biophysical and molecular mechanisms of auditory hair cell mechanotransduction.Nat Commun. 2011 Nov 1;2:523. doi: 10.1038/ncomms1533. Nat Commun. 2011. PMID: 22045002 Free PMC article. Review.
-
Genetic loci associated with skin pigmentation in African Americans and their effects on vitamin D deficiency.PLoS Genet. 2021 Feb 18;17(2):e1009319. doi: 10.1371/journal.pgen.1009319. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33600456 Free PMC article.
-
Quantitative trait loci on chromosome 5 for susceptibility to frequency-specific effects on hearing in DBA/2J mice.Exp Anim. 2015;64(3):241-51. doi: 10.1538/expanim.14-0110. Epub 2015 Mar 10. Exp Anim. 2015. PMID: 25765874 Free PMC article.
-
Discovery Genetics - The History and Future of Spontaneous Mutation Research.Curr Protoc Mouse Biol. 2012 Jun 1;2:103-118. doi: 10.1002/9780470942390.mo110200. Curr Protoc Mouse Biol. 2012. PMID: 25364627 Free PMC article.
-
Homozygosity mapping reveals mutations of GRXCR1 as a cause of autosomal-recessive nonsyndromic hearing impairment.Am J Hum Genet. 2010 Feb 12;86(2):138-47. doi: 10.1016/j.ajhg.2009.12.017. Epub 2010 Feb 4. Am J Hum Genet. 2010. PMID: 20137778 Free PMC article.
References
-
- Lim D.J., Anniko M. Developmental morphology of the mouse inner ear. A scanning electron microscopic observation. Acta Otolaryngol. 1985;422(Supplement):1–69. - PubMed
-
- Goodyear R.J., Marcotti W., Kros C.J., Richardson G.P. Development and properties of stereociliary link types in hair cells of the mouse cochlea. J. Comp. Neurol. 2005;485:75–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R29 DC003049/DC/NIDCD NIH HHS/United States
- R29 DC03049/DC/NIDCD NIH HHS/United States
- P30-DC05188/DC/NIDCD NIH HHS/United States
- R01-DC002842/DC/NIDCD NIH HHS/United States
- Z01 DC000048/ImNIH/Intramural NIH HHS/United States
- P30 DC005188/DC/NIDCD NIH HHS/United States
- DC62108/DC/NIDCD NIH HHS/United States
- R01 DC002842/DC/NIDCD NIH HHS/United States
- R01-DC003049/DC/NIDCD NIH HHS/United States
- R01 DC004314/DC/NIDCD NIH HHS/United States
- R01 DC003049/DC/NIDCD NIH HHS/United States
- T32-DC000011/DC/NIDCD NIH HHS/United States
- R01-DC004314/DC/NIDCD NIH HHS/United States
- R01 DC004301/DC/NIDCD NIH HHS/United States
- T32 DC000011/DC/NIDCD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
