

Human rhinovirus 14 enters rhabdomyosarcoma cells expressing icam-1 by a clathrin-, caveolin-, and flotillin-independent pathway
- PMID: 20130060
- PMCID: PMC2849509
- DOI: 10.1128/JVI.01693-09
Human rhinovirus 14 enters rhabdomyosarcoma cells expressing icam-1 by a clathrin-, caveolin-, and flotillin-independent pathway
Abstract
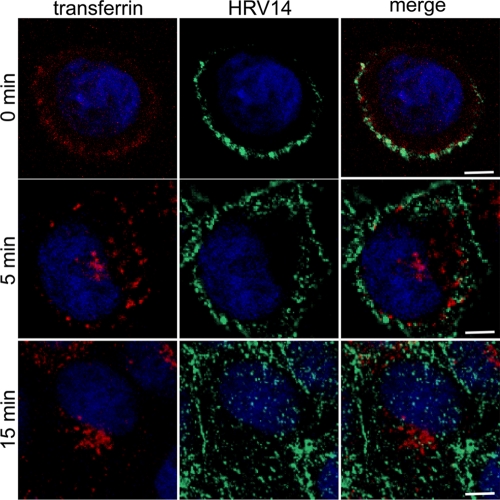
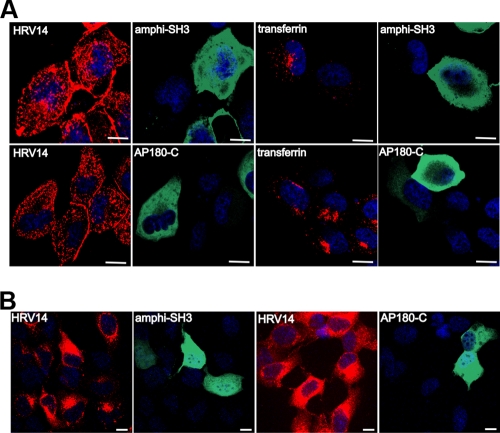
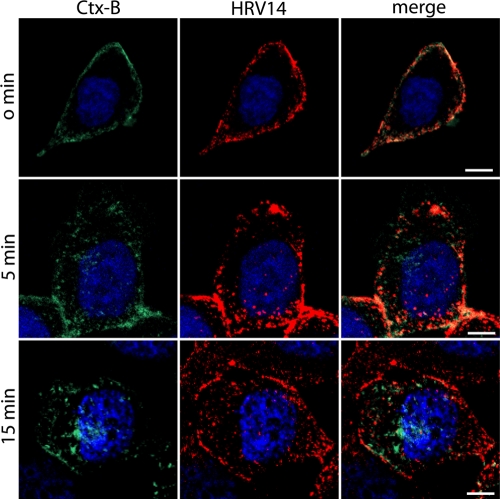
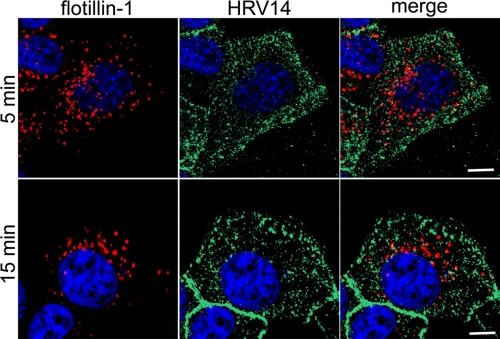
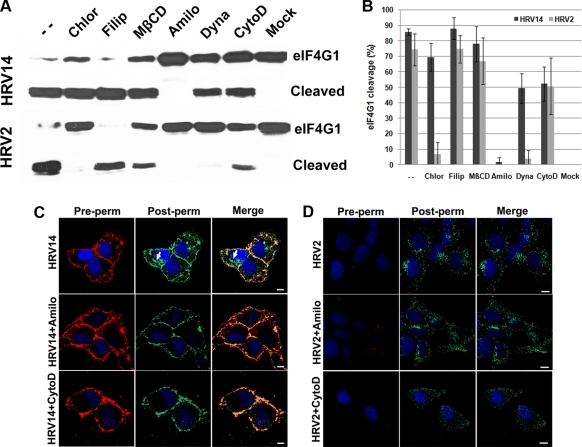
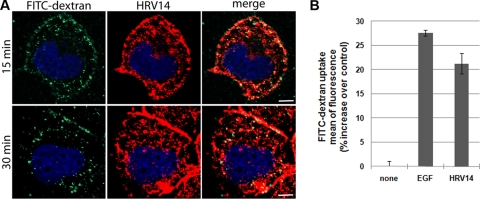
Intercellular adhesion molecule 1 (ICAM-1) mediates binding and entry of major group human rhinoviruses (HRVs). Whereas the entry pathway of minor group HRVs has been studied in detail and is comparatively well understood, the pathway taken by major group HRVs is largely unknown. Use of immunofluorescence microscopy, colocalization with specific endocytic markers, dominant negative mutants, and pharmacological inhibitors allowed us to demonstrate that the major group virus HRV14 enters rhabdomyosarcoma cells transfected to express human ICAM-1 in a clathrin-, caveolin-, and flotillin-independent manner. Electron microscopy revealed that many virions accumulated in long tubular structures, easily distinguishable from clathrin-coated pits and caveolae. Virus entry was strongly sensitive to the Na(+)/H(+) ion exchange inhibitor amiloride and moderately sensitive to cytochalasin D. Thus, cellular uptake of HRV14 occurs via a pathway exhibiting some, but not all, characteristics of macropinocytosis and is similar to that recently described for adenovirus 3 entry via alpha(v) integrin/CD46 in HeLa cells.
Figures

Similar articles
-
Entry of a heparan sulphate-binding HRV8 variant strictly depends on dynamin but not on clathrin, caveolin, and flotillin.Virology. 2011 Mar 30;412(1):55-67. doi: 10.1016/j.virol.2010.12.042. Epub 2011 Jan 22. Virology. 2011. PMID: 21262518
-
Entry of human rhinovirus 89 via ICAM-1 into HeLa epithelial cells is inhibited by actin skeleton disruption and by bafilomycin.Arch Virol. 2014 Jan;159(1):125-40. doi: 10.1007/s00705-013-1797-1. Epub 2013 Aug 4. Arch Virol. 2014. PMID: 23913188
-
Internalization of human rhinovirus 14 into HeLa and ICAM-1-transfected BHK cells.Med Microbiol Immunol. 1997 Jun;186(1):1-9. doi: 10.1007/s004300050039. Med Microbiol Immunol. 1997. PMID: 9255760
-
Uncoating of human rhinoviruses.Rev Med Virol. 2010 Sep;20(5):281-97. doi: 10.1002/rmv.654. Rev Med Virol. 2010. PMID: 20629045 Review.
-
ICAM-1 receptors and cold viruses.Pharm Acta Helv. 2000 Mar;74(2-3):291-7. doi: 10.1016/s0031-6865(99)00056-4. Pharm Acta Helv. 2000. PMID: 10812972 Free PMC article. Review.
Cited by
-
Productive entry pathways of human rhinoviruses.Adv Virol. 2012;2012:826301. doi: 10.1155/2012/826301. Epub 2012 Nov 26. Adv Virol. 2012. PMID: 23227049 Free PMC article.
-
Productive Entry of Foot-and-Mouth Disease Virus via Macropinocytosis Independent of Phosphatidylinositol 3-Kinase.Sci Rep. 2016 Jan 13;6:19294. doi: 10.1038/srep19294. Sci Rep. 2016. PMID: 26757826 Free PMC article.
-
Vaccine and Wild-Type Strains of Yellow Fever Virus Engage Distinct Entry Mechanisms and Differentially Stimulate Antiviral Immune Responses.mBio. 2016 Feb 9;7(1):e01956-15. doi: 10.1128/mBio.01956-15. mBio. 2016. PMID: 26861019 Free PMC article.
-
Endosome rupture enables enteroviruses from the family Picornaviridae to infect cells.Commun Biol. 2024 Nov 8;7(1):1465. doi: 10.1038/s42003-024-07147-9. Commun Biol. 2024. PMID: 39511383 Free PMC article.
-
Different rotavirus strains enter MA104 cells through different endocytic pathways: the role of clathrin-mediated endocytosis.J Virol. 2010 Sep;84(18):9161-9. doi: 10.1128/JVI.00731-10. Epub 2010 Jul 14. J Virol. 2010. PMID: 20631149 Free PMC article.
References
-
- Bayer, N., E. Prchla, M. Schwab, D. Blaas, and R. Fuchs. 1999. Human rhinovirus HRV14 uncoats from early endosomes in the presence of bafilomycin. FEBS Lett. 463:175-178. - PubMed
-
- Bayer, N., D. Schober, M. Huttinger, D. Blaas, and R. Fuchs. 2001. Inhibition of clathrin-dependent endocytosis has multiple effects on human rhinovirus serotype 2 cell entry. J. Biol. Chem. 276:3952-3962. - PubMed
-
- Bonazzi, M., S. Spano, G. Turacchio, C. Cericola, C. Valente, A. Colanzi, H. S. Kweon, V. W. Hsu, E. V. Polishchuck, R. S. Polishchuck, M. Sallese, T. Pulvirenti, D. Corda, and A. Luini. 2005. CtBP3/BARS drives membrane fission in dynamin-independent transport pathways. Nat. Cell Biol. 7:570-580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
