

Mucosal parainfluenza virus-vectored vaccine against Ebola virus replicates in the respiratory tract of vector-immune monkeys and is immunogenic
- PMID: 20129638
- PMCID: PMC2842940
- DOI: 10.1016/j.virol.2010.01.015
Mucosal parainfluenza virus-vectored vaccine against Ebola virus replicates in the respiratory tract of vector-immune monkeys and is immunogenic
Abstract
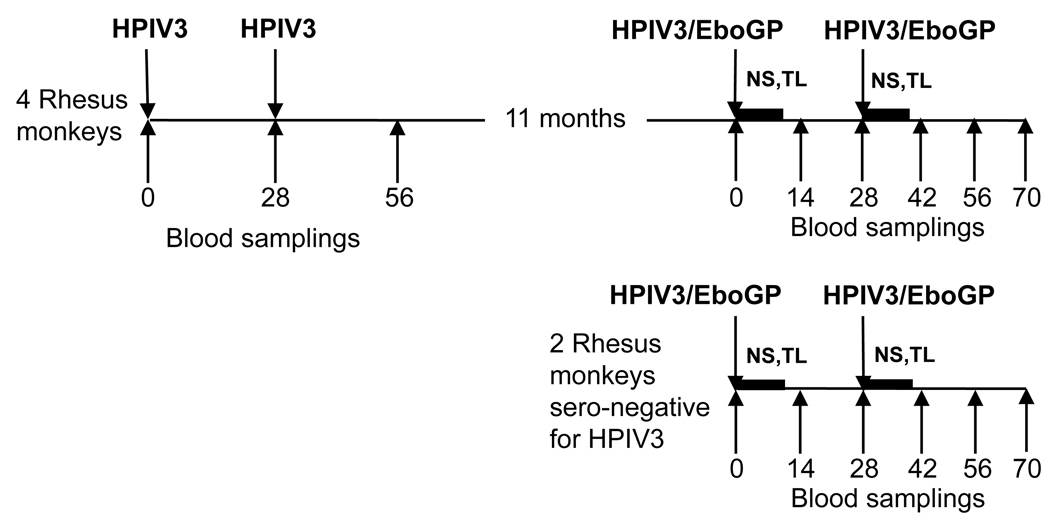
We previously used human parainfluenza virus type 3 (HPIV3) as a vector to express the Ebola virus (EBOV) GP glycoprotein. The resulting HPIV3/EboGP vaccine was immunogenic and protective against EBOV challenge in a non-human primate model. However, it remained unclear whether the vaccine would be effective in adults due to preexisting immunity to HPIV3. Here, the immunogenicity of HPIV3/EboGP was compared in HPIV3-naive and HPIV3-immune Rhesus monkeys. After a single dose of HPIV3/EboGP, the titers of EBOV-specific serum ELISA or neutralization antibodies were substantially less in HPIV3-immune animals compared to HPIV3-naive animals. However, after two doses, which were previously determined to be required for complete protection against EBOV challenge, the antibody titers were indistinguishable between the two groups. The vaccine virus appeared to replicate, at a reduced level, in the respiratory tract despite the preexisting immunity. This may reflect the known ability of HPIV3 to re-infect and may also reflect the presence of EBOV GP in the vector virion, which confers resistance to neutralization in vitro by HPIV3-specific antibodies. These data suggest that HPIV3/EboGP will be immunogenic in adults as well as children.
Published by Elsevier Inc.
Figures
Similar articles
-
Respiratory tract immunization of non-human primates with a Newcastle disease virus-vectored vaccine candidate against Ebola virus elicits a neutralizing antibody response.Vaccine. 2010 Dec 10;29(1):17-25. doi: 10.1016/j.vaccine.2010.10.024. Epub 2010 Oct 27. Vaccine. 2010. PMID: 21034822 Free PMC article.
-
A paramyxovirus-vectored intranasal vaccine against Ebola virus is immunogenic in vector-immune animals.Virology. 2008 Aug 1;377(2):255-64. doi: 10.1016/j.virol.2008.04.029. Virology. 2008. PMID: 18570964 Free PMC article.
-
Attenuated Human Parainfluenza Virus Type 1 Expressing Ebola Virus Glycoprotein GP Administered Intranasally Is Immunogenic in African Green Monkeys.J Virol. 2017 Apr 28;91(10):e02469-16. doi: 10.1128/JVI.02469-16. Print 2017 May 15. J Virol. 2017. PMID: 28250127 Free PMC article.
-
The Ebola virus glycoprotein and its immune responses across multiple vaccine platforms.Expert Rev Vaccines. 2020 Mar;19(3):267-277. doi: 10.1080/14760584.2020.1738225. Epub 2020 Mar 16. Expert Rev Vaccines. 2020. PMID: 32129120 Review.
-
Correlates of vaccine-induced protective immunity against Ebola virus disease.Semin Immunol. 2018 Oct;39:65-72. doi: 10.1016/j.smim.2018.07.003. Epub 2018 Jul 21. Semin Immunol. 2018. PMID: 30041831 Review.
Cited by
-
Antibody quality and protection from lethal Ebola virus challenge in nonhuman primates immunized with rabies virus based bivalent vaccine.PLoS Pathog. 2013;9(5):e1003389. doi: 10.1371/journal.ppat.1003389. Epub 2013 May 30. PLoS Pathog. 2013. PMID: 23737747 Free PMC article.
-
A single intranasal dose of human parainfluenza virus type 3-vectored vaccine induces effective antibody and memory T cell response in the lungs and protects hamsters against SARS-CoV-2.NPJ Vaccines. 2022 Apr 25;7(1):47. doi: 10.1038/s41541-022-00471-3. NPJ Vaccines. 2022. PMID: 35468973 Free PMC article.
-
Viral vectored vaccines: design, development, preventive and therapeutic applications in human diseases.Signal Transduct Target Ther. 2023 Apr 7;8(1):149. doi: 10.1038/s41392-023-01408-5. Signal Transduct Target Ther. 2023. PMID: 37029123 Free PMC article. Review.
-
Ebola: Lessons on Vaccine Development.Annu Rev Microbiol. 2018 Sep 8;72:423-446. doi: 10.1146/annurev-micro-090817-062414. Annu Rev Microbiol. 2018. PMID: 30200851 Free PMC article. Review.
-
Progress in filovirus vaccine development: evaluating the potential for clinical use.Expert Rev Vaccines. 2011 Jan;10(1):63-77. doi: 10.1586/erv.10.152. Expert Rev Vaccines. 2011. PMID: 21162622 Free PMC article. Review.
References
-
- Barouch DH, Pau MG, Custers JH, Koudstaal W, Kostense S, Havenga MJ, Truitt DM, Sumida SM, Kishko MG, Arthur JC, Korioth-Schmitz B, Newberg MH, Gorgone DA, Lifton MA, Panicali DL, Nabel GJ, Letvin NL, Goudsmit J. Immunogenicity of recombinant adenovirus serotype 35 vaccine in the presence of pre-existing anti-Ad5 immunity. J Immunol. 2004;172(10):6290–6297. - PubMed
-
- Bukreyev A, Collins PL. Filovirus vaccines: what challenges are left? Expert Rev Vaccines. 2010;9(1):5–8. - PubMed
-
- Bukreyev A, Lamirande EW, Buchholz UJ, Vogel LN, Elkins WR, St Claire M, Murphy BR, Subbarao K, Collins PL. Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS. Lancet. 2004;363(9427):2122–2127. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
