

Architecture of the RNA polymerase II-TFIIF complex revealed by cross-linking and mass spectrometry
- PMID: 20094031
- PMCID: PMC2810376
- DOI: 10.1038/emboj.2009.401
Architecture of the RNA polymerase II-TFIIF complex revealed by cross-linking and mass spectrometry
Abstract
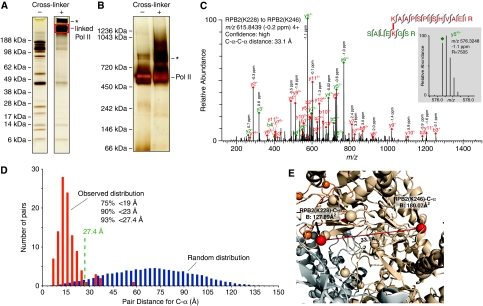
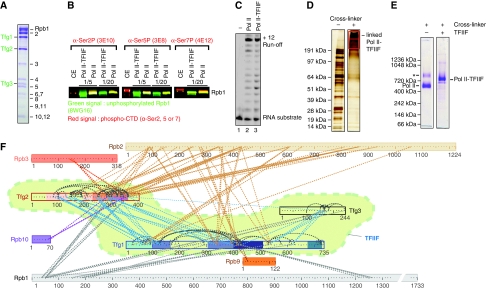
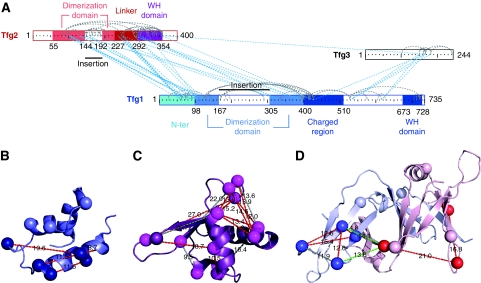
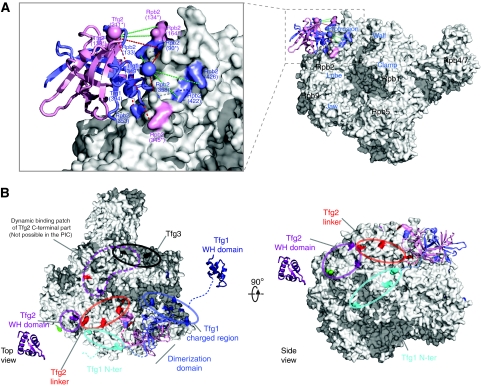
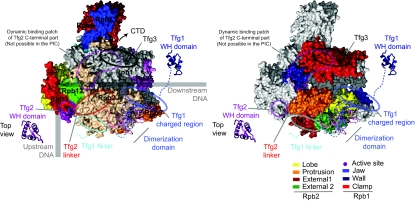
Higher-order multi-protein complexes such as RNA polymerase II (Pol II) complexes with transcription initiation factors are often not amenable to X-ray structure determination. Here, we show that protein cross-linking coupled to mass spectrometry (MS) has now sufficiently advanced as a tool to extend the Pol II structure to a 15-subunit, 670 kDa complex of Pol II with the initiation factor TFIIF at peptide resolution. The N-terminal regions of TFIIF subunits Tfg1 and Tfg2 form a dimerization domain that binds the Pol II lobe on the Rpb2 side of the active centre cleft near downstream DNA. The C-terminal winged helix (WH) domains of Tfg1 and Tfg2 are mobile, but the Tfg2 WH domain can reside at the Pol II protrusion near the predicted path of upstream DNA in the initiation complex. The linkers between the dimerization domain and the WH domains in Tfg1 and Tfg2 are located to the jaws and protrusion, respectively. The results suggest how TFIIF suppresses non-specific DNA binding and how it helps to recruit promoter DNA and to set the transcription start site. This work establishes cross-linking/MS as an integrated structure analysis tool for large multi-protein complexes.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Position of the general transcription factor TFIIF within the RNA polymerase II transcription preinitiation complex.EMBO J. 2010 Feb 17;29(4):706-16. doi: 10.1038/emboj.2009.386. Epub 2009 Dec 24. EMBO J. 2010. PMID: 20033062 Free PMC article.
-
Conserved architecture of the core RNA polymerase II initiation complex.Nat Commun. 2014 Jul 10;5:4310. doi: 10.1038/ncomms5310. Nat Commun. 2014. PMID: 25007739
-
Structural and binding studies of the C-terminal domains of yeast TFIIF subunits Tfg1 and Tfg2.Proteins. 2012 Feb;80(2):519-29. doi: 10.1002/prot.23217. Epub 2011 Nov 17. Proteins. 2012. PMID: 22095626
-
RNA polymerase II elongation factors use conserved regulatory mechanisms.Curr Opin Struct Biol. 2024 Feb;84:102766. doi: 10.1016/j.sbi.2023.102766. Epub 2024 Jan 4. Curr Opin Struct Biol. 2024. PMID: 38181687 Review.
-
Conservation between the RNA polymerase I, II, and III transcription initiation machineries.Mol Cell. 2012 Feb 24;45(4):439-46. doi: 10.1016/j.molcel.2012.01.023. Mol Cell. 2012. PMID: 22365827 Review.
Cited by
-
The Importance of Non-accessible Crosslinks and Solvent Accessible Surface Distance in Modeling Proteins with Restraints From Crosslinking Mass Spectrometry.Mol Cell Proteomics. 2016 Jul;15(7):2491-500. doi: 10.1074/mcp.M116.058560. Epub 2016 May 5. Mol Cell Proteomics. 2016. PMID: 27150526 Free PMC article.
-
Ska3 Ensures Timely Mitotic Progression by Interacting Directly With Microtubules and Ska1 Microtubule Binding Domain.Sci Rep. 2016 Sep 26;6:34042. doi: 10.1038/srep34042. Sci Rep. 2016. PMID: 27667719 Free PMC article.
-
Cytoplasmic TAF2-TAF8-TAF10 complex provides evidence for nuclear holo-TFIID assembly from preformed submodules.Nat Commun. 2015 Jan 14;6:6011. doi: 10.1038/ncomms7011. Nat Commun. 2015. PMID: 25586196 Free PMC article.
-
Uncoupling Promoter Opening from Start-Site Scanning.Mol Cell. 2015 Jul 2;59(1):133-8. doi: 10.1016/j.molcel.2015.05.021. Epub 2015 Jun 11. Mol Cell. 2015. PMID: 26073544 Free PMC article.
-
Histone Interaction Landscapes Visualized by Crosslinking Mass Spectrometry in Intact Cell Nuclei.Mol Cell Proteomics. 2018 Oct;17(10):2018-2033. doi: 10.1074/mcp.RA118.000924. Epub 2018 Jul 18. Mol Cell Proteomics. 2018. PMID: 30021884 Free PMC article.
References
-
- Armache KJ, Mitterweger S, Meinhart A, Cramer P (2005) Structures of complete RNA polymerase II and its subcomplex, Rpb4/7. J Biol Chem 280: 7131–7134 - PubMed
-
- Brueckner F, Hennecke U, Carell T, Cramer P (2007) CPD damage recognition by transcribing RNA polymerase II. Science 315: 859–862 - PubMed
-
- Chafin DR, Claussen TJ, Price DH (1991) Identification and purification of a yeast protein that affects elongation by RNA polymerase II. J Biol Chem 266: 9256–9262 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
