

Alternative splicing modulates Ubx protein function in Drosophila melanogaster
- PMID: 20038634
- PMCID: PMC2845342
- DOI: 10.1534/genetics.109.112086
Alternative splicing modulates Ubx protein function in Drosophila melanogaster
Abstract
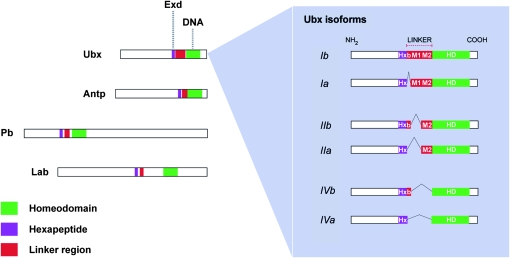
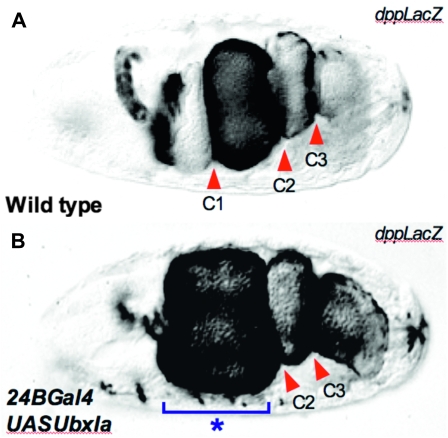
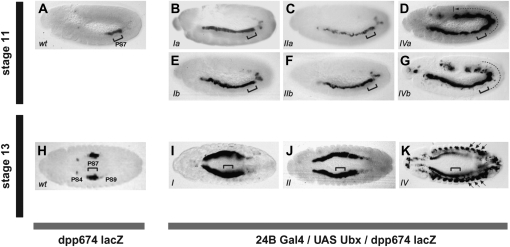
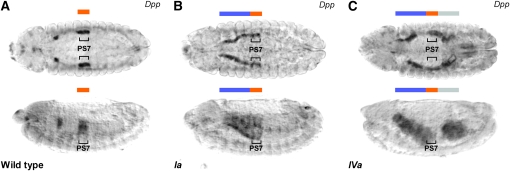
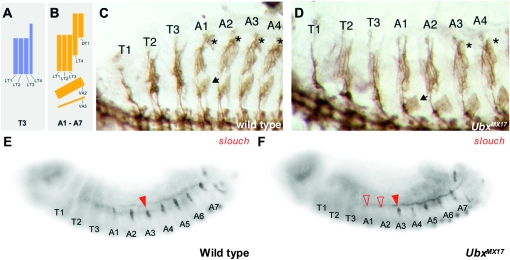
The Drosophila Hox gene Ultrabithorax (Ubx) produces a family of protein isoforms through alternative splicing. Isoforms differ from one another by the presence of optional segments-encoded by individual exons-that modify the distance between the homeodomain and a cofactor-interaction module termed the "YPWM" motif. To investigate the functional implications of Ubx alternative splicing, here we analyze the in vivo effects of the individual Ubx isoforms on the activation of a natural Ubx molecular target, the decapentaplegic (dpp) gene, within the embryonic mesoderm. These experiments show that the Ubx isoforms differ in their abilities to activate dpp in mesodermal tissues during embryogenesis. Furthermore, using a Ubx mutant that reduces the full Ubx protein repertoire to just one single isoform, we obtain specific anomalies affecting the patterning of anterior abdominal muscles, demonstrating that Ubx isoforms are not functionally interchangeable during embryonic mesoderm development. Finally, a series of experiments in vitro reveals that Ubx isoforms also vary in their capacity to bind DNA in presence of the cofactor Extradenticle (Exd). Altogether, our results indicate that the structural changes produced by alternative splicing have functional implications for Ubx protein function in vivo and in vitro. Since other Hox genes also produce splicing isoforms affecting similar protein domains, we suggest that alternative splicing may represent an underestimated regulatory system modulating Hox gene specificity during fly development.
Figures

Similar articles
-
Integration of RNA processing and expression level control modulates the function of the Drosophila Hox gene Ultrabithorax during adult development.Development. 2011 Jan;138(1):107-16. doi: 10.1242/dev.051409. Epub 2010 Nov 29. Development. 2011. PMID: 21115609
-
Distinct functions of homeodomain-containing and homeodomain-less isoforms encoded by homothorax.Genes Dev. 2006 Jun 15;20(12):1636-50. doi: 10.1101/gad.1412606. Genes Dev. 2006. PMID: 16778079 Free PMC article.
-
Evolutionary conservation of the structure and expression of alternatively spliced Ultrabithorax isoforms from Drosophila.Genetics. 1994 Mar;136(3):965-77. doi: 10.1093/genetics/136.3.965. Genetics. 1994. PMID: 7911773 Free PMC article.
-
Roles for intrinsic disorder and fuzziness in generating context-specific function in Ultrabithorax, a Hox transcription factor.Adv Exp Med Biol. 2012;725:86-105. doi: 10.1007/978-1-4614-0659-4_6. Adv Exp Med Biol. 2012. PMID: 22399320 Review.
-
Ultrabithorax and the evolution of insect forewing/hindwing differentiation.Curr Opin Insect Sci. 2017 Feb;19:8-15. doi: 10.1016/j.cois.2016.10.007. Epub 2016 Nov 2. Curr Opin Insect Sci. 2017. PMID: 28521947 Review.
Cited by
-
Regulated functional alternative splicing in Drosophila.Nucleic Acids Res. 2012 Jan;40(1):1-10. doi: 10.1093/nar/gkr648. Epub 2011 Sep 8. Nucleic Acids Res. 2012. PMID: 21908400 Free PMC article. Review.
-
Onychophoran Hox genes and the evolution of arthropod Hox gene expression.Front Zool. 2014 Mar 5;11(1):22. doi: 10.1186/1742-9994-11-22. Front Zool. 2014. PMID: 24594097 Free PMC article.
-
Control of Hox transcription factor concentration and cell-to-cell variability by an auto-regulatory switch.Development. 2019 Jan 25;146(12):dev168179. doi: 10.1242/dev.168179. Development. 2019. PMID: 30642837 Free PMC article.
-
The intrinsically disordered regions of the Drosophila melanogaster Hox protein ultrabithorax select interacting proteins based on partner topology.PLoS One. 2014 Oct 6;9(10):e108217. doi: 10.1371/journal.pone.0108217. eCollection 2014. PLoS One. 2014. PMID: 25286318 Free PMC article.
-
Improving Hox protein classification across the major model organisms.PLoS One. 2010 May 25;5(5):e10820. doi: 10.1371/journal.pone.0010820. PLoS One. 2010. PMID: 20520839 Free PMC article.
References
-
- Abu-Shaar, M., and R. S. Mann, 1998. Generation of multiple antagonistic domains along the proximodistal axis during Drosophila leg development. Development 125 3821–3830. - PubMed
-
- Akam, M., 1998. Hox genes, homeosis and the evolution of segment identity: no need for hopeless monsters. Int. J. Dev. Biol. 42 445–451. - PubMed
-
- Akam, M. E., A. Martinez-Arias, R. Weinzierl and C. D. Wilde, 1985. Function and expression of ultrabithorax in the Drosophila embryo. Cold Spring Harbor Symp. Quant. Biol. 50 195–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
