

Review
doi: 10.1038/nrc2681.
Emerging roles of ATF2 and the dynamic AP1 network in cancer
Affiliations
- PMID: 20029425
- PMCID: PMC2874064
- DOI: 10.1038/nrc2681
Item in Clipboard
Review
Emerging roles of ATF2 and the dynamic AP1 network in cancer
Nat Rev Cancer.
2010 Jan.
Erratum in
- Nat Rev Cancer. 2010 May;10(5):379
Abstract
Cooperation among transcription factors is central for their ability to execute specific transcriptional programmes. The AP1 complex exemplifies a network of transcription factors that function in unison under normal circumstances and during the course of tumour development and progression. This Perspective summarizes our current understanding of the changes in members of the AP1 complex and the role of ATF2 as part of this complex in tumorigenesis.
Conflict of interest statement
Figures
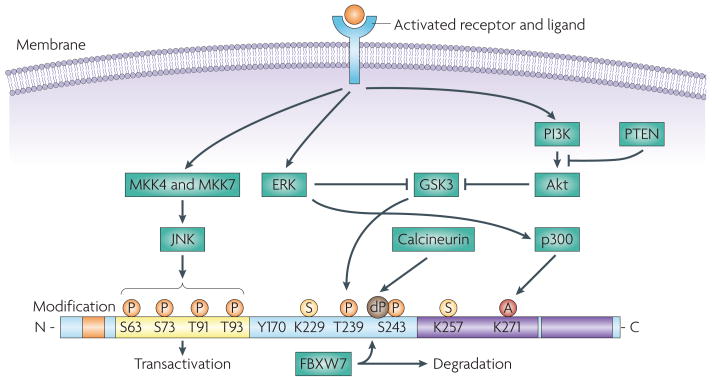
JUN is encoded by a 3.34 kb intronless gene, located on chromosome 1 (1p32-p31) and results in the expression of a 334 amino acid protein product composed of four main domains, which are involved in DNA binding, transcription and dimerization. JUN activity is regulated by post-translational modifications, which are largely controlled by components of the MAPK family of serine and threonine kinases, including JUN N-terminal kinase (JNK), ERK and p38 isoforms. JUN is phosphorylated on Ser63 and Ser73 by JNK, increasing its stability and transactivation potential. JNK also phosphorylates Thr91 and Thr93, which are required for DNA binding and activation of its transcriptional activity. JUN is subject to ubiquitylation and this requires phosphorylation at Thr239 by glycogen synthase kinase 3 (GSK3). GSK3 can target JUN only once Ser243 is phosphorylated. Phosphorylation on these sites is required for recruitment of the F-box and WD domain repeated 7 (FBXW7) ubiquitin ligase. Inactivation of GSK3, owing to the activation of ERK and PI3K–Akt signalling cascades results in JUN stabilization,. The effect of GSK3 can be antagonized by the dephosphorylation of Ser243 by calcineurin. JUN can be sumoylated on Lys257 and Lys229, which leads to a reduced transcriptional activity. ERK induces the acetylation of the lysine residues in the JUN DNA binding region, thereby increasing JUN transcriptional activity. Post-translational modifications are indicated as small coloured circles. The four domains are indicated as follows: the δ-domain is orange, the basic region (DNA binding) is blue, the transactivation domain is yellow and the leucine zipper is purple. A, acetylation; dP, dephosphorylation; P, phosphorylation; S, sumoylation.
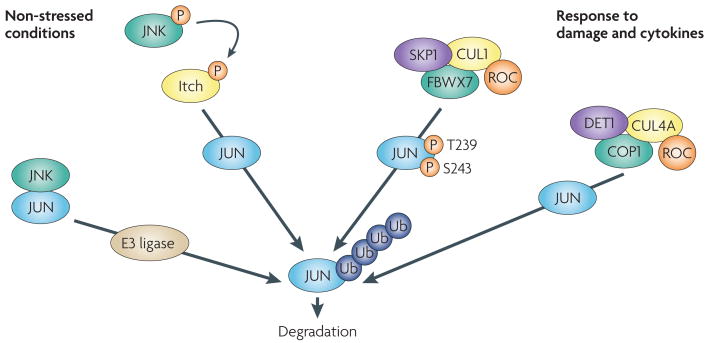
Several mechanisms exist to limit JUN stablity. Under non-stressed conditions, JUN N-terminal kinase (JNK) is tightly bound to JUN, targeting JUN for ubiquitylation and degradation. Activation of JNK by stress results in JUN phosphorylation and dissociation from JNK, enhancing its protein stability, although phosphorylation on Ser63 and Ser73 promotes the F-box and WD domain repeated 7 (FBXW7)-mediated degradation of JUN. As part of a feedforward mechanism, JNK also phosphorylates the E3 ligase Itch in T cells after stimulation, accelerating degradation of JUN and JUNB, independently of Ser63 and Ser73 phosphorylation. Phosphorylation on JUN at Thr239 by glycogen synthase kinase 3 (GsK3) (FIG. 1) allows FBXW7 binding and ubiquitin-mediated degradation by a Skp1-Cullin1-F-box (SCF) complex. Following osmotic stress, MEKK1 can activate JUN N-terminal phosphorylation by activating MKK4, an upstream kinase for JNKs that can also function as a JUN E3 ubiquitin ligase, promoting its ubiquitin–proteasome-dependent degradation (not shown). De-etiolated 1 (DET1) contributes to JUN degradation by promoting the formation of a ubiquitin ligase complex containing DNA damage binding protein 1 (DDB1), cullin 4A (CUL4A), regulator of cullins 1 (ROC1) and constitutively photomorphogenic 1 (COP1). Other mechanisms modulating JUN ubiquitin-mediated degradation have been reported. P, phosphorylation; Ub, ubiquitylation.
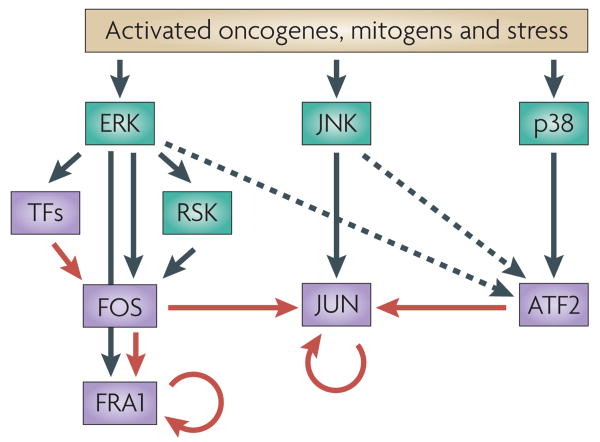
ERK, JUN N-terminal kinase (JNK) and p38 are predominantly responsible for the phosphorylation and activation of FOS, JUN and ATF2 respectively (black arrows) in response to stress, mitogens or oncogene activation. ERK and JNK also regulate FOS and JUN degradation, respectively (not depicted) and participate in ATF2 activation (dashed arrows). ERK also induces the transcription of FOS through the activation of non-AP1 transcription factors (TFs). Transcription of some AP1 proteins (that is, JUN and FRA1) is regulated by crosstalk among AP1 complexes (red arrows) as well as by autoregulation (circular red arrows).
Similar articles
-
[Ap1-like cis-elements in 5'-regulatory region of human apolipoprotein A-I gene].Mol Biol (Mosk). 2008 Mar-Apr;42(2):295-305. Mol Biol (Mosk). 2008. PMID: 18610838 Russian.
-
Transient receptor potential melastatin-3 (TRPM3)-induced activation of AP-1 requires Ca2+ ions and the transcription factors c-Jun, ATF2, and ternary complex factor.Mol Pharmacol. 2015 Apr;87(4):617-28. doi: 10.1124/mol.114.095695. Epub 2015 Jan 9. Mol Pharmacol. 2015. PMID: 25576487
-
Mutual regulation of c-Jun and ATF2 by transcriptional activation and subcellular localization.EMBO J. 2006 Mar 8;25(5):1058-69. doi: 10.1038/sj.emboj.7601020. Epub 2006 Mar 2. EMBO J. 2006. PMID: 16511568 Free PMC article.
-
ATF2, a paradigm of the multifaceted regulation of transcription factors in biology and disease.Pharmacol Res. 2017 May;119:347-357. doi: 10.1016/j.phrs.2017.02.004. Epub 2017 Feb 15. Pharmacol Res. 2017. PMID: 28212892 Free PMC article. Review.
-
The roles of ATF2 (activating transcription factor 2) in tumorigenesis.Biochem Soc Trans. 2012 Feb;40(1):230-4. doi: 10.1042/BST20110630. Biochem Soc Trans. 2012. PMID: 22260696 Review.
Cited by
-
FOSL1 promotes metastasis of head and neck squamous cell carcinoma through super-enhancer-driven transcription program.Mol Ther. 2021 Aug 4;29(8):2583-2600. doi: 10.1016/j.ymthe.2021.03.024. Epub 2021 Mar 29. Mol Ther. 2021. PMID: 33794365 Free PMC article.
-
Emerging roles of activating transcription factor 2 in the development of breast cancer: a comprehensive review.Precis Clin Med. 2023 Oct 25;6(4):pbad028. doi: 10.1093/pcmedi/pbad028. eCollection 2023 Dec. Precis Clin Med. 2023. PMID: 37955015 Free PMC article. Review.
-
Rough set soft computing cancer classification and network: one stone, two birds.Cancer Inform. 2010 Jul 15;9:139-45. doi: 10.4137/cin.s4874. Cancer Inform. 2010. PMID: 20706619 Free PMC article.
-
The potential of activator protein 1 (AP-1) in cancer targeted therapy.Front Immunol. 2023 Jul 6;14:1224892. doi: 10.3389/fimmu.2023.1224892. eCollection 2023. Front Immunol. 2023. PMID: 37483616 Free PMC article. Review.
-
Syk: a new target for attenuation of Helicobacter pylori-induced gastric mucosal inflammatory responses.Inflammopharmacology. 2019 Apr;27(2):203-211. doi: 10.1007/s10787-019-00577-6. Epub 2019 Feb 28. Inflammopharmacology. 2019. PMID: 30820719 Review.
References
-
- Lee W, Mitchell P, Tjian R. Purified transcription factor AP1 interacts with TPA-inducible enhancer elements. Cell. 1987;49:741–752. - PubMed
-
- Angel P, et al. Phorbol ester-inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell. 1987;49:729–739. - PubMed
-
- Wisdom R. AP1: One switch for many signals. Exp Cell Res. 1999;253:180–185. - PubMed
-
- Eferl R, Wagner EF. AP1: a double-edged sword in tumorigenesis. Nature Rev Cancer. 2003;3:859–868. - PubMed
-
- Angel P, Karin M. The role of Jun, Fos and the AP1 complex in cell-proliferation and transformation. Biochem Biophys Acta. 1991;1072:129–157. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- CA051995/CA/NCI NIH HHS/United States
- R01 CA059908-12/CA/NCI NIH HHS/United States
- R01 CA051995/CA/NCI NIH HHS/United States
- CA117927/CA/NCI NIH HHS/United States
- R01 CA099961/CA/NCI NIH HHS/United States
- R01 CA099961-06A2/CA/NCI NIH HHS/United States
- R01 CA117927/CA/NCI NIH HHS/United States
- R01 CA059908/CA/NCI NIH HHS/United States
- CA099961/CA/NCI NIH HHS/United States
- T32 CA121929/CA/NCI NIH HHS/United States
- R01 CA117927-04/CA/NCI NIH HHS/United States
- R01 CA051995-18/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
