

Asymmetric deactivation of HIV-1 gp41 following fusion inhibitor binding
- PMID: 19956769
- PMCID: PMC2776349
- DOI: 10.1371/journal.ppat.1000674
Asymmetric deactivation of HIV-1 gp41 following fusion inhibitor binding
Abstract
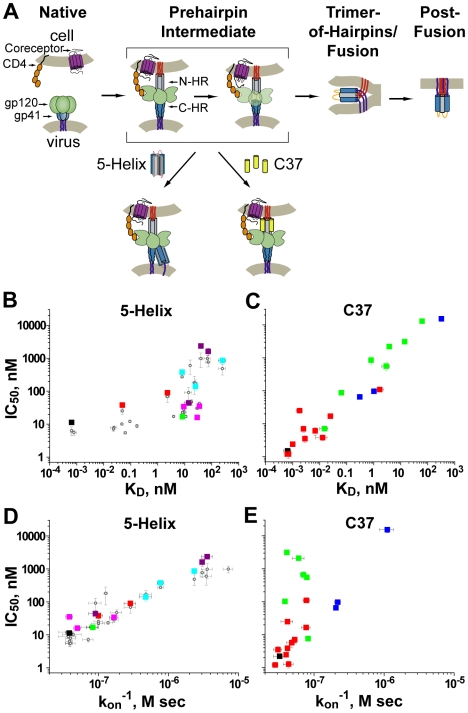
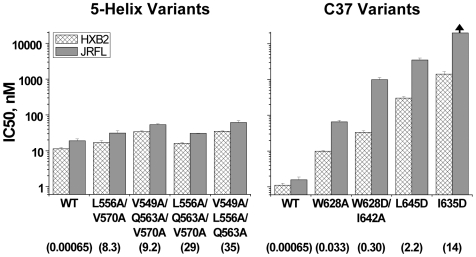
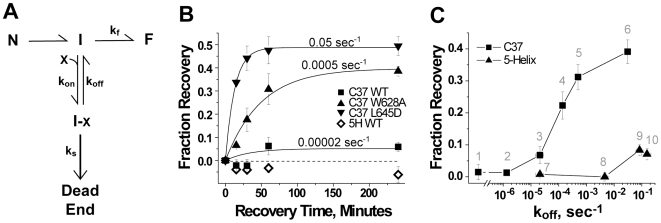
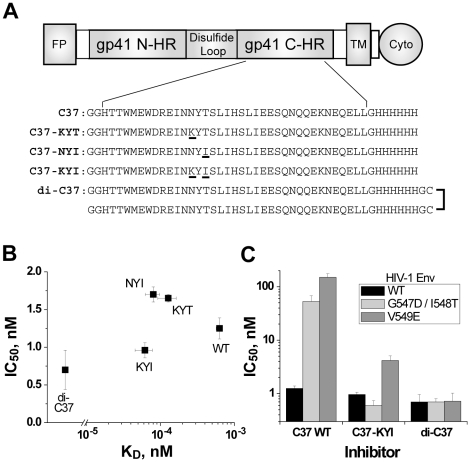
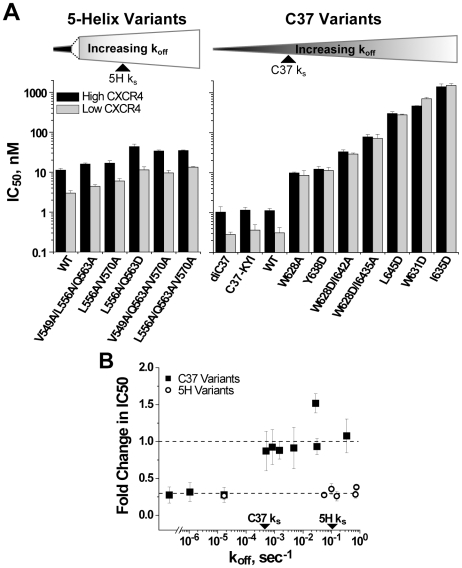
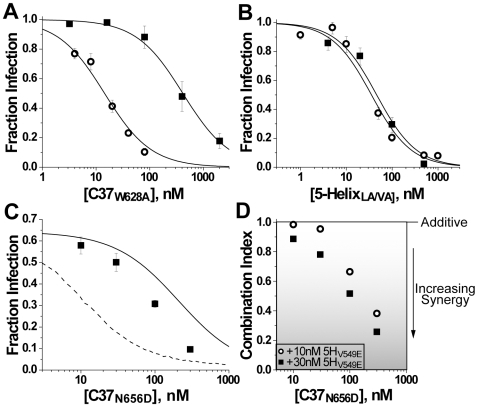
Both equilibrium and nonequilibrium factors influence the efficacy of pharmaceutical agents that target intermediate states of biochemical reactions. We explored the intermediate state inhibition of gp41, part of the HIV-1 envelope glycoprotein complex (Env) that promotes viral entry through membrane fusion. This process involves a series of gp41 conformational changes coordinated by Env interactions with cellular CD4 and a chemokine receptor. In a kinetic window between CD4 binding and membrane fusion, the N- and C-terminal regions of the gp41 ectodomain become transiently susceptible to inhibitors that disrupt Env structural transitions. In this study, we sought to identify kinetic parameters that influence the antiviral potency of two such gp41 inhibitors, C37 and 5-Helix. Employing a series of C37 and 5-Helix variants, we investigated the physical properties of gp41 inhibition, including the ability of inhibitor-bound gp41 to recover its fusion activity once inhibitor was removed from solution. Our results indicated that antiviral activity critically depended upon irreversible deactivation of inhibitor-bound gp41. For C37, which targets the N-terminal region of the gp41 ectodomain, deactivation was a slow process that depended on chemokine receptor binding to Env. For 5-Helix, which targets the C-terminal region of the gp41 ectodomain, deactivation occurred rapidly following inhibitor binding and was independent of chemokine receptor levels. Due to this kinetic disparity, C37 inhibition was largely reversible, while 5-Helix inhibition was functionally irreversible. The fundamental difference in deactivation mechanism points to an unappreciated asymmetry in gp41 following inhibitor binding and impacts the development of improved fusion inhibitors and HIV-1 vaccines. The results also demonstrate how the activities of intermediate state inhibitors critically depend upon the final disposition of inhibitor-bound states.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
HIV-1 gp41 Residues Modulate CD4-Induced Conformational Changes in the Envelope Glycoprotein and Evolution of a Relaxed Conformation of gp120.J Virol. 2018 Jul 31;92(16):e00583-18. doi: 10.1128/JVI.00583-18. Print 2018 Aug 15. J Virol. 2018. PMID: 29875245 Free PMC article.
-
Conserved Residue Asn-145 in the C-Terminal Heptad Repeat Region of HIV-1 gp41 is Critical for Viral Fusion and Regulates the Antiviral Activity of Fusion Inhibitors.Viruses. 2019 Jul 3;11(7):609. doi: 10.3390/v11070609. Viruses. 2019. PMID: 31277353 Free PMC article.
-
Interactions of HIV-1 inhibitory peptide T20 with the gp41 N-HR coiled coil.J Biol Chem. 2009 Feb 6;284(6):3619-27. doi: 10.1074/jbc.M809269200. Epub 2008 Dec 10. J Biol Chem. 2009. PMID: 19073602 Free PMC article.
-
HIV-1 gp41: mediator of fusion and target for inhibition.AIDS Rev. 2003 Oct-Dec;5(4):214-21. AIDS Rev. 2003. PMID: 15012000 Review.
-
Membrane-transferring regions of gp41 as targets for HIV-1 fusion inhibition and viral neutralization.Curr Top Med Chem. 2011 Dec;11(24):2985-96. doi: 10.2174/156802611798808460. Curr Top Med Chem. 2011. PMID: 22044227 Review.
Cited by
-
Conformational Stabilization of Gp41-Mimetic Miniproteins Opens Up New Ways of Inhibiting HIV-1 Fusion.Int J Mol Sci. 2022 Mar 3;23(5):2794. doi: 10.3390/ijms23052794. Int J Mol Sci. 2022. PMID: 35269936 Free PMC article.
-
Selection with a peptide fusion inhibitor corresponding to the first heptad repeat of HIV-1 gp41 identifies two genetic pathways conferring cross-resistance to peptide fusion inhibitors corresponding to the first and second heptad repeats (HR1 and HR2) of gp41.J Virol. 2011 Dec;85(24):12929-38. doi: 10.1128/JVI.05391-11. Epub 2011 Oct 12. J Virol. 2011. PMID: 21994458 Free PMC article.
-
Characterization of resistance to a potent D-peptide HIV entry inhibitor.Retrovirology. 2019 Oct 22;16(1):28. doi: 10.1186/s12977-019-0489-7. Retrovirology. 2019. PMID: 31640718 Free PMC article.
-
Structural basis of HIV-1 neutralization by affinity matured Fabs directed against the internal trimeric coiled-coil of gp41.PLoS Pathog. 2010 Nov 11;6(11):e1001182. doi: 10.1371/journal.ppat.1001182. PLoS Pathog. 2010. PMID: 21085615 Free PMC article.
-
Complex interplay of kinetic factors governs the synergistic properties of HIV-1 entry inhibitors.J Biol Chem. 2017 Oct 6;292(40):16498-16510. doi: 10.1074/jbc.M117.791731. Epub 2017 Jul 10. J Biol Chem. 2017. PMID: 28696261 Free PMC article.
References
-
- Robertson JG. Mechanistic basis of enzyme-targeted drugs. Biochemistry. 2005;44:5561–5571. - PubMed
-
- Lundqvist T, Fisher SL, Kern G, Folmer RH, Xue Y, et al. Exploitation of structural and regulatory diversity in glutamate racemases. Nature. 2007;447:817–822. - PubMed
-
- Steger HK, Root MJ. Kinetic dependence to HIV-1 entry inhibition. J Biol Chem. 2006;281:25813–25821. - PubMed
-
- Root MJ, Steger HK. HIV-1 gp41 as a target for viral entry inhibition. Curr Pharm Des. 2004;10:1805–1825. - PubMed
-
- Chan DC, Fass D, Berger JM, Kim PS. Core structure of gp41 from the HIV envelope glycoprotein. Cell. 1997;89:263–273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
