

Identification and characterization of a small molecule inhibitor of formin-mediated actin assembly
- PMID: 19942139
- PMCID: PMC2784894
- DOI: 10.1016/j.chembiol.2009.10.006
Identification and characterization of a small molecule inhibitor of formin-mediated actin assembly
Abstract
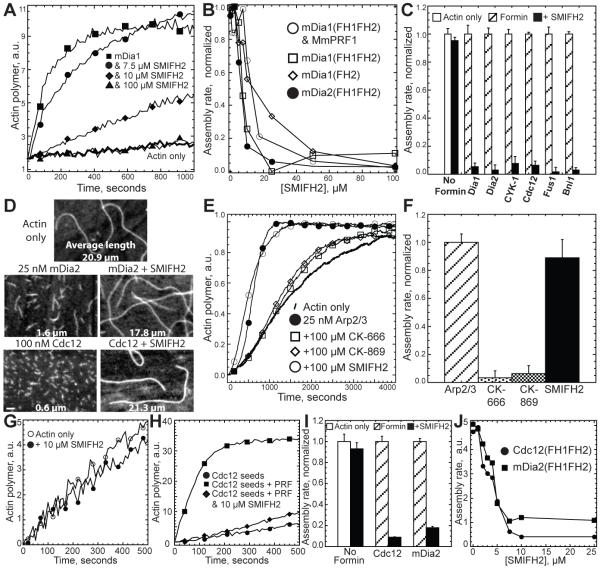
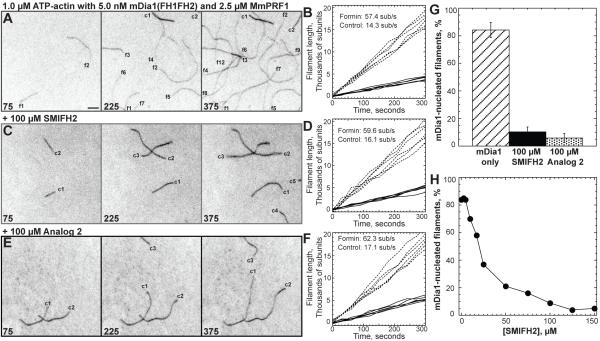
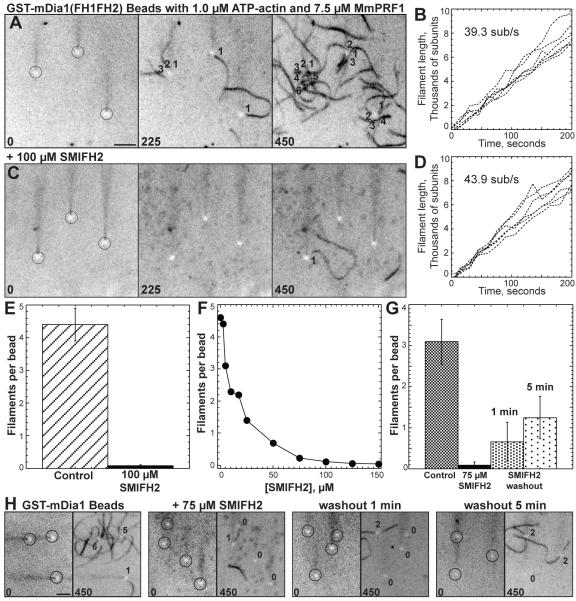
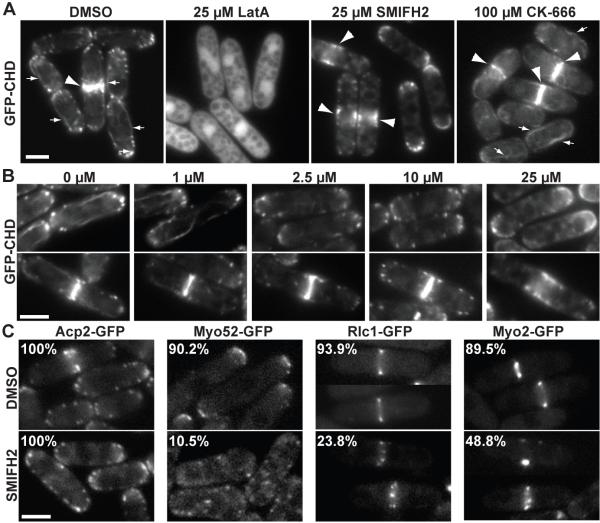
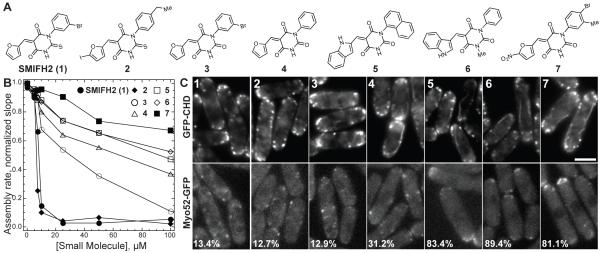
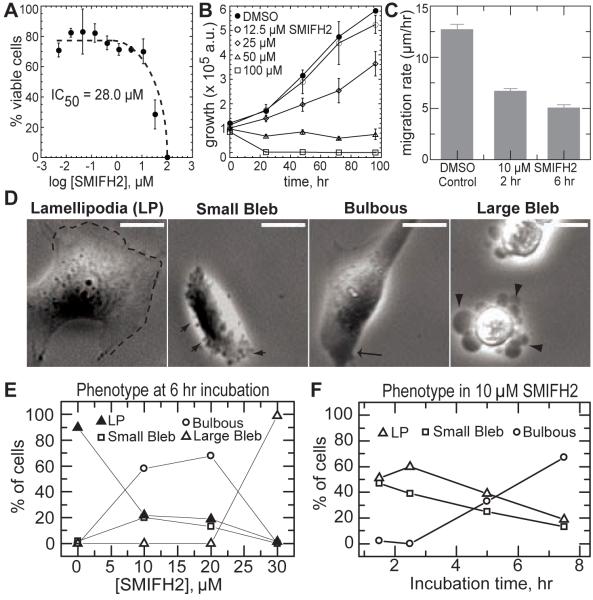
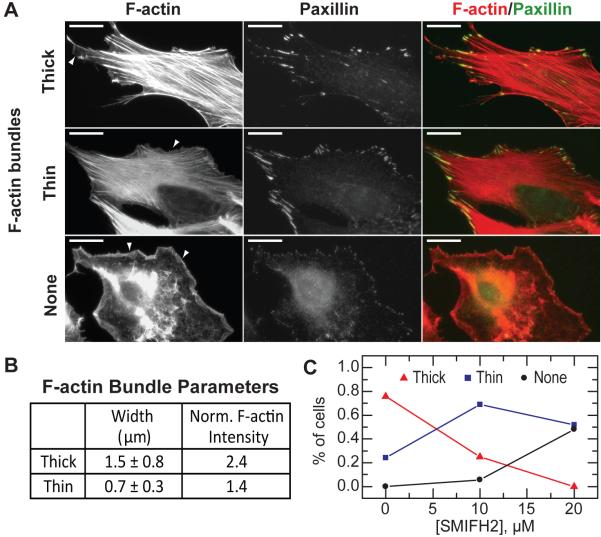
Formins stimulate actin filament assembly for fundamental cellular processes including division, adhesion, establishing polarity, and motility. A formin inhibitor would be useful because most cells express multiple formins whose functions are not known and because metastatic tumor formation depends on the deregulation of formin-dependent processes. We identified a general small molecule inhibitor of formin homology 2 domains (SMIFH2) by screening compounds for the ability to prevent formin-mediated actin assembly in vitro. SMIFH2 targets formins from evolutionarily diverse organisms including yeast, nematode worm, and mice, with a half-maximal inhibitor concentration of approximately 5 to 15 microM. SMIFH2 prevents both formin nucleation and processive barbed end elongation and decreases formin's affinity for the barbed end. Furthermore, low micromolar concentrations of SMIFH2 disrupt formin-dependent, but not Arp2/3 complex-dependent, actin cytoskeletal structures in fission yeast and mammalian NIH 3T3 fibroblasts.
Figures
Comment in
-
Inhibitors target actin nucleators.Chem Biol. 2009 Nov 25;16(11):1125-6. doi: 10.1016/j.chembiol.2009.11.001. Chem Biol. 2009. PMID: 19942132
Similar articles
-
Inhibitors target actin nucleators.Chem Biol. 2009 Nov 25;16(11):1125-6. doi: 10.1016/j.chembiol.2009.11.001. Chem Biol. 2009. PMID: 19942132
-
Alterations to the broad-spectrum formin inhibitor SMIFH2 modulate potency but not specificity.Sci Rep. 2022 Aug 8;12(1):13520. doi: 10.1038/s41598-022-17685-z. Sci Rep. 2022. PMID: 35941181 Free PMC article.
-
SMIFH2 has effects on Formins and p53 that perturb the cell cytoskeleton.Sci Rep. 2015 Apr 30;5:9802. doi: 10.1038/srep09802. Sci Rep. 2015. PMID: 25925024 Free PMC article.
-
Investigating Mammalian Formins with SMIFH2 Fifteen Years in: Novel Targets and Unexpected Biology.Int J Mol Sci. 2023 May 21;24(10):9058. doi: 10.3390/ijms24109058. Int J Mol Sci. 2023. PMID: 37240404 Free PMC article. Review.
-
Review of the mechanism of processive actin filament elongation by formins.Cell Motil Cytoskeleton. 2009 Aug;66(8):606-17. doi: 10.1002/cm.20379. Cell Motil Cytoskeleton. 2009. PMID: 19459187 Free PMC article. Review.
Cited by
-
Negative control of cytokinesis by stress-activated MAPK signaling.Curr Genet. 2021 Oct;67(5):715-721. doi: 10.1007/s00294-021-01155-6. Epub 2021 Mar 31. Curr Genet. 2021. PMID: 33791858 Review.
-
AtFH1 formin mutation affects actin filament and microtubule dynamics in Arabidopsis thaliana.J Exp Bot. 2013 Jan;64(2):585-97. doi: 10.1093/jxb/ers351. Epub 2012 Dec 1. J Exp Bot. 2013. PMID: 23202131 Free PMC article.
-
Formin-mediated actin polymerization at cell-cell junctions stabilizes E-cadherin and maintains monolayer integrity during wound repair.Mol Biol Cell. 2016 Sep 15;27(18):2844-56. doi: 10.1091/mbc.E16-06-0429. Epub 2016 Jul 20. Mol Biol Cell. 2016. PMID: 27440924 Free PMC article.
-
Actin networks modulate heterogeneous NF-κB dynamics in response to TNFα.Elife. 2024 Aug 7;13:e86042. doi: 10.7554/eLife.86042. Elife. 2024. PMID: 39110005 Free PMC article.
-
Formin-generated actomyosin arcs propel T cell receptor microcluster movement at the immune synapse.J Cell Biol. 2016 Nov 7;215(3):383-399. doi: 10.1083/jcb.201603080. Epub 2016 Oct 31. J Cell Biol. 2016. PMID: 27799367 Free PMC article.
References
-
- Chhabra ES, Higgs HN. The many faces of actin: matching assembly factors with cellular structures. Nat Cell Biol. 2007;9:1110–1121. - PubMed
-
- Eisenmann KM, West RA, Hildebrand D, Kitchen SM, Peng J, Sigler R, Zhang J, Siminovitch KA, Alberts AS. T cell responses in mammalian diaphanous-related formin mDia1 knock-out mice. J Biol chem. 2007;282:25152–25158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
