

Alternative processing of primary microRNA transcripts by Drosha generates 5' end variation of mature microRNA
- PMID: 19859542
- PMCID: PMC2762519
- DOI: 10.1371/journal.pone.0007566
Alternative processing of primary microRNA transcripts by Drosha generates 5' end variation of mature microRNA
Abstract
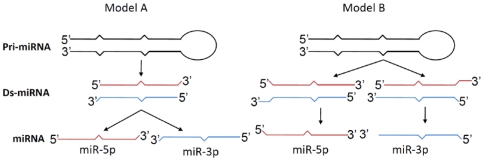
Background: It is generally believed that the miRNA processing machinery ensures the generation of a mature miRNA with a fixed sequence, particularly at its 5' end. However, we and others have recently noted that the ends of a given mature miRNA are not absolutely fixed, but subject to variation. Neither the significance nor the mechanism behind the generation of such miRNA polymorphism is understood. miR-142 is an abundantly expressed miRNA in hematopoietic cells and exhibits a high frequency of 5' end polymorphism.
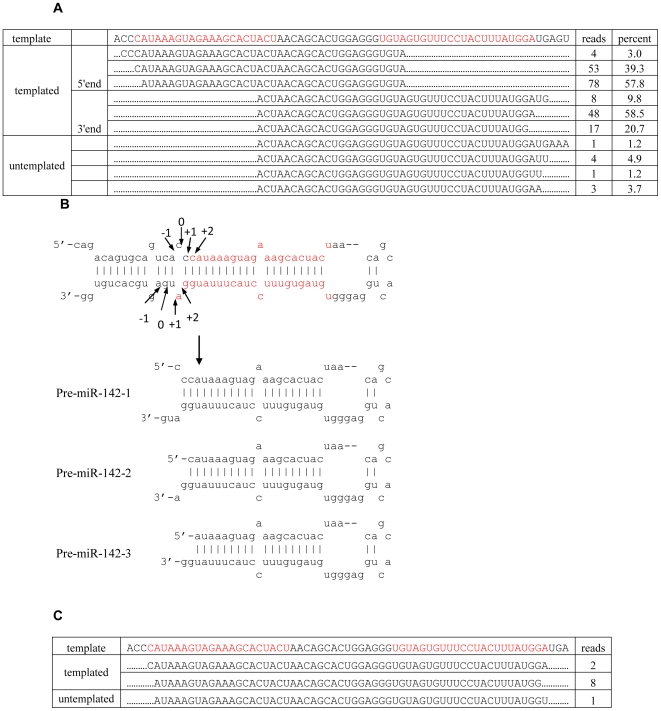
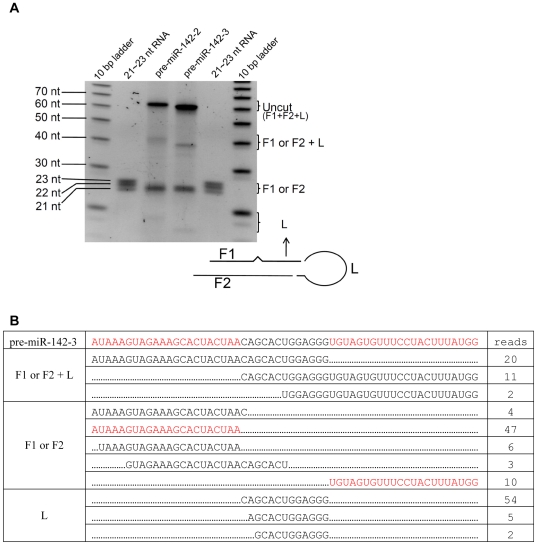
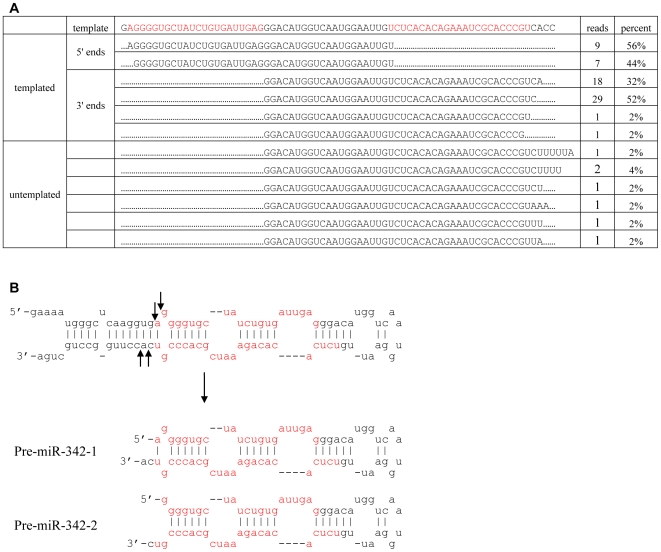
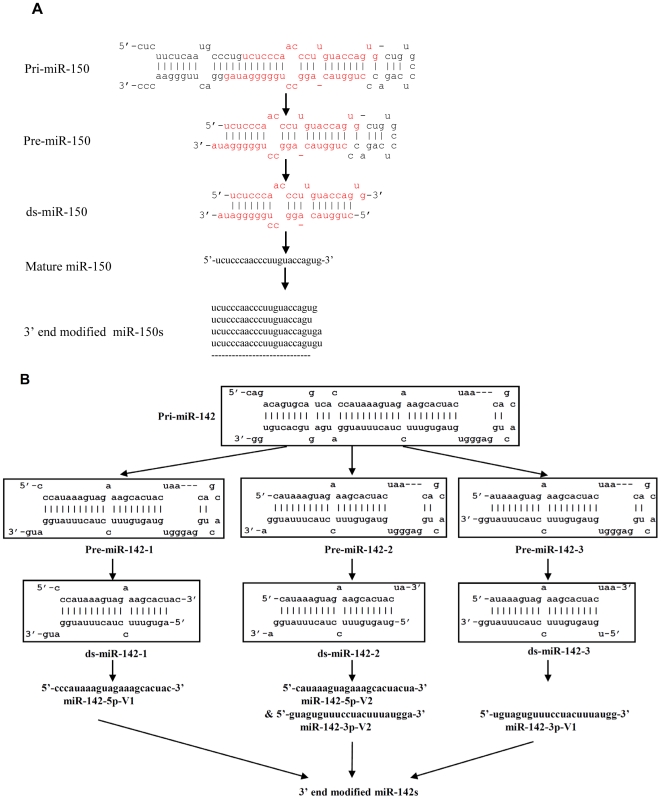
Methodology/principal findings: Here we show that a shift in the Drosha processing of pri-miRNA generates multiple forms of miR-142s in vivo with differing 5' ends that might target different genes. Sequence analysis of several pre-miRNA ends cloned from T cells reveals that unlike many other pri-miRNAs that are processed into a single pre-miRNA, pri-miR-142 is processed into 3 distinct pre-miR-142s. Dicer processing studies suggest that each of the 3 pre-miR-142s is processed into a distinct double-stranded miRNA, giving rise to 4 mature miRNA variants that might regulate different target gene pools.
Conclusions/significance: Thus, alternative Drosha processing might be a novel mechanism for diversification of the miRNA target gene pool.
Conflict of interest statement
Figures

Similar articles
-
A central role for the primary microRNA stem in guiding the position and efficiency of Drosha processing of a viral pri-miRNA.RNA. 2014 Jul;20(7):1068-77. doi: 10.1261/rna.044537.114. Epub 2014 May 22. RNA. 2014. PMID: 24854622 Free PMC article.
-
Re-evaluation of the roles of DROSHA, Export in 5, and DICER in microRNA biogenesis.Proc Natl Acad Sci U S A. 2016 Mar 29;113(13):E1881-9. doi: 10.1073/pnas.1602532113. Epub 2016 Mar 14. Proc Natl Acad Sci U S A. 2016. PMID: 26976605 Free PMC article.
-
Structural Differences between Pri-miRNA Paralogs Promote Alternative Drosha Cleavage and Expand Target Repertoires.Cell Rep. 2019 Jan 8;26(2):447-459.e4. doi: 10.1016/j.celrep.2018.12.054. Cell Rep. 2019. PMID: 30625327 Free PMC article.
-
MicroRNA biogenesis: isolation and characterization of the microprocessor complex.Methods Mol Biol. 2006;342:33-47. doi: 10.1385/1-59745-123-1:33. Methods Mol Biol. 2006. PMID: 16957365 Review.
-
New insights into pri-miRNA processing and accumulation in plants.Wiley Interdiscip Rev RNA. 2015 Sep-Oct;6(5):533-45. doi: 10.1002/wrna.1292. Epub 2015 Jun 29. Wiley Interdiscip Rev RNA. 2015. PMID: 26119101 Review.
Cited by
-
Northern blotting analysis of microRNAs, their precursors and RNA interference triggers.BMC Mol Biol. 2011 Apr 11;12:14. doi: 10.1186/1471-2199-12-14. BMC Mol Biol. 2011. PMID: 21481235 Free PMC article.
-
Mismatched and wobble base pairs govern primary microRNA processing by human Microprocessor.Nat Commun. 2020 Apr 21;11(1):1926. doi: 10.1038/s41467-020-15674-2. Nat Commun. 2020. PMID: 32317642 Free PMC article.
-
A meta-analysis revealed insights into the sources, conservation and impact of microRNA 5'-isoforms in four model species.Nucleic Acids Res. 2014 Feb;42(3):1427-41. doi: 10.1093/nar/gkt967. Epub 2013 Oct 30. Nucleic Acids Res. 2014. PMID: 24178030 Free PMC article.
-
Effective tools for RNA-derived therapeutics: siRNA interference or miRNA mimicry.Theranostics. 2021 Aug 11;11(18):8771-8796. doi: 10.7150/thno.62642. eCollection 2021. Theranostics. 2021. PMID: 34522211 Free PMC article. Review.
-
MicroRNA-focused CRISPR/Cas9 screen identifies miR-142 as a key regulator of Epstein-Barr virus reactivation.PLoS Pathog. 2024 Jun 17;20(6):e1011970. doi: 10.1371/journal.ppat.1011970. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38885264 Free PMC article.
References
-
- Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Bernstein E, Kim SY, Carmell MA, Murchison EP, Alcorn H, et al. Dicer is essential for mouse development. Nat Genet. 2003;35:215–217. - PubMed
-
- Garzon R, Calin GA, Croce CM. MicroRNAs in Cancer. Annu Rev Med. 2009;60:167–179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
