

Mycobacterial Esx-3 is required for mycobactin-mediated iron acquisition
- PMID: 19846780
- PMCID: PMC2774023
- DOI: 10.1073/pnas.0900589106
Mycobacterial Esx-3 is required for mycobactin-mediated iron acquisition
Abstract
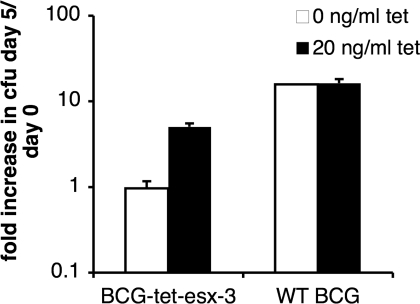
The Esx secretion pathway is conserved across Gram-positive bacteria. Esx-1, the best-characterized system, is required for virulence of Mycobacterium tuberculosis, although its precise function during infection remains unclear. Esx-3, a paralogous system present in all mycobacterial species, is required for growth in vitro. Here, we demonstrate that mycobacteria lacking Esx-3 are defective in acquiring iron. To compete for the limited iron available in the host and the environment, these organisms use mycobactin, high-affinity iron-binding molecules. In the absence of Esx-3, mycobacteria synthesize mycobactin but are unable to use the bound iron and are impaired severely for growth during macrophage infection. Mycobacteria thus require a specialized secretion system for acquiring iron from siderophores.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Separable roles for Mycobacterium tuberculosis ESX-3 effectors in iron acquisition and virulence.Proc Natl Acad Sci U S A. 2016 Jan 19;113(3):E348-57. doi: 10.1073/pnas.1523321113. Epub 2016 Jan 4. Proc Natl Acad Sci U S A. 2016. PMID: 26729876 Free PMC article.
-
Mycobacterial Esx-3 requires multiple components for iron acquisition.mBio. 2014 May 6;5(3):e01073-14. doi: 10.1128/mBio.01073-14. mBio. 2014. PMID: 24803520 Free PMC article.
-
Inability to detect mycobactin in mycobacteria-infected tissues suggests an alternative iron acquisition mechanism by mycobacteria in vivo.Microb Pathog. 1993 Mar;14(3):229-38. doi: 10.1006/mpat.1993.1022. Microb Pathog. 1993. PMID: 8321124
-
Iron acquisition strategies in mycobacteria.Tuberculosis (Edinb). 2015 Mar;95(2):123-30. doi: 10.1016/j.tube.2015.01.004. Epub 2015 Jan 17. Tuberculosis (Edinb). 2015. PMID: 25636179 Review.
-
ESX-3 secretion system in Mycobacterium: An overview.Biochimie. 2024 Jan;216:46-55. doi: 10.1016/j.biochi.2023.10.013. Epub 2023 Oct 23. Biochimie. 2024. PMID: 37879428 Review.
Cited by
-
In Silico Drug Repurposing Studies for the Discovery of Novel Salicyl-AMP Ligase (MbtA)Inhibitors.Pathogens. 2023 Dec 9;12(12):1433. doi: 10.3390/pathogens12121433. Pathogens. 2023. PMID: 38133316 Free PMC article.
-
Essential Role of the ESX-5 Secretion System in Outer Membrane Permeability of Pathogenic Mycobacteria.PLoS Genet. 2015 May 4;11(5):e1005190. doi: 10.1371/journal.pgen.1005190. eCollection 2015 May. PLoS Genet. 2015. PMID: 25938982 Free PMC article.
-
Insight into the biology of Mycobacterium mucogenicum and Mycobacterium neoaurum clade members.Sci Rep. 2019 Dec 17;9(1):19259. doi: 10.1038/s41598-019-55464-5. Sci Rep. 2019. PMID: 31848383 Free PMC article.
-
The Rich Tapestry of Bacterial Protein Translocation Systems.Protein J. 2019 Aug;38(4):389-408. doi: 10.1007/s10930-019-09862-3. Protein J. 2019. PMID: 31407127 Free PMC article. Review.
-
Genome-wide Phenotypic Profiling Identifies and Categorizes Genes Required for Mycobacterial Low Iron Fitness.Sci Rep. 2019 Aug 6;9(1):11394. doi: 10.1038/s41598-019-47905-y. Sci Rep. 2019. PMID: 31388080 Free PMC article.
References
-
- DiGiuseppe Champion PA, Cox JS. Protein secretion systems in Mycobacteria. Cell Microbiol. 2007;9:1376–1384. - PubMed
-
- Abdallah AM, et al. Type VII secretion—Mycobacteria show the way. Nat Rev Microbiol. 2007;5:883–891. - PubMed
-
- Cole ST, et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature. 1998;393:537–544. - PubMed
-
- Tekaia F, et al. Analysis of the proteome of Mycobacterium tuberculosis in silico. Tuber Lung Dis. 1999;79:329–342. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
