

Review
doi: 10.1038/nchembio.250.
Targeting proteins for degradation
Affiliations
- PMID: 19841631
- PMCID: PMC4228941
- DOI: 10.1038/nchembio.250
Item in Clipboard
Review
Targeting proteins for degradation
Nat Chem Biol.
2009 Nov.
Erratum in
- Nat Chem Biol. 2009 Dec;5(12):954
Abstract
Protein degradation plays a central role in many cellular functions. Misfolded and damaged proteins are removed from the cell to avoid toxicity. The concentrations of regulatory proteins are adjusted by degradation at the appropriate time. Both foreign and native proteins are digested into small peptides as part of the adaptive immune response. In eukaryotic cells, an ATP-dependent protease called the proteasome is responsible for much of this proteolysis. Proteins are targeted for proteasomal degradation by a two-part degron, which consists of a proteasome binding signal and a degradation initiation site. Here we describe how both components contribute to the specificity of degradation.
Figures
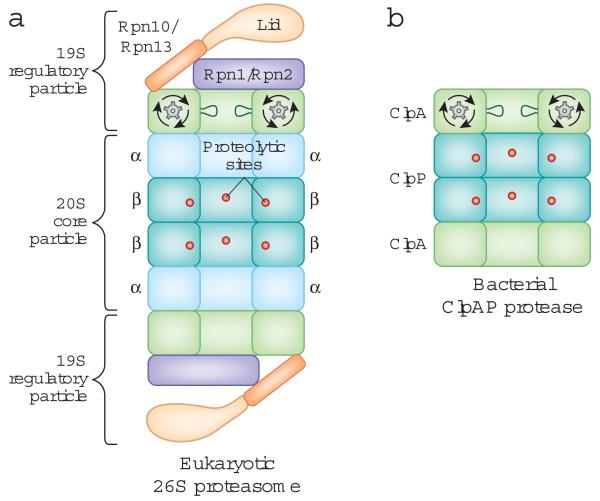
The overall structure of the eukaryotic proteasome and the bacterial ClpAP protease. The two proteases have a common architecture. The protease sites are buried in an internal chamber of the core particle, which is capped by regulatory particles that control access to the proteolytic chamber and contain an ATPase hexameric ring. A: Side-on cross-section of the eukaryotic 26S proteasome. The 20S core particle is flanked by 19S regulatory particles, and the proteolytic sites are located in the β-rings of the 20S core particle. The scaffold proteins Rpn1 and Rpn2, the ubiquitin receptors Rpn10 and Rpn13, and the loops lining the ATPase ring are depicted. Only one set of loops is shown for clarity. B: Side-on cross-section of the ClpAP protease from Escherichia coli. ClpP contains the proteolytic sites and is capped on both ends by the ClpA ATPase ring.

Mechanisms of targeting ubiquitinated proteins to the proteasome. A: Some substrates bind directly to the 19S regulatory particle subunits Rpn10 or Rpn13. B: Ubiquitinated substrates can also be carried to the proteasome by adaptor proteins, which bind both to the ubiquitin modification of the substrate and to the proteasome. C: Several proteins are targeted to the proteasome independent of ubiquitination. Two cases are shown. On the left, a protein is engaged by the proteasome in the absence of any separate proteasome-binding motif. This might occur if the initiation site has a particularly high affinity for the site on the proteasome that engages substrates. On the right, ornithine decarboxylase (ODC) is shown as it is degraded by the proteasome when bound to antizyme 1 (AZ1). AZ1 binding to ODC exposes a 37 amino acid proteasome recognition signal in the C-terminus of ODC, where degradation then begins. The 37 amino acid signal may function as a high affinity proteasome binding site by itself, or AZ1 may act like an adaptor protein.

An initiation site is required for protein degradation. A: A protein lacking an initiation site escapes degradation. The proteasome-targeted protein does not have an effective initiation site. The deubiquitinating enzymes trim the ubiquitin chain before the proteasome engages the substrate fully and initiates degradation. Thus, the intact substrate is released from the proteasome. B: A protein containing an effective initiation site is degraded by the proteasome. A ubiquitinated protein is recognized by the proteasome, and the ubiquitin ligase Hul5 and the deubiquitinating enzymes Ubp6 and Uch37 extend or trim the ubiquitin chain, respectively. The proteasome engages the protein at its unstructured region, which leads to unfolding and translocation of the polypeptide into the degradation chamber. In this case, the proteasome engages the substrate so rapidly that the deubiquitinating enzymes are not able to remove enough ubiquitin moieties to cause substrate release.

The Cdc48 / p97 targeting machine. Cdc48 / p97 is an ATPase ring complex involved in a range of cellular processes. It can serve as a scaffold for the assembly of E3 and E4 ligases, deubiquitinating enzymes, and ubiquitinated substrates. The fate of a substrate may be determined by the various enzymatic activities associated with Cdc48. A: The Ufd1-Npl4 heterodimer recruits a polyubiquitinated substrate to Cdc48. The polyubiquitin chain on the substrate is lengthened by Ufd2, which has E4 activity. The substrate is then passed from Ufd2 to the adaptor protein Rad23 (or Dsk2), which shuttles the protein to the proteasome for degradation. Figure adapted from. B: Cdc48-associated deubiquitinating enzymes (DUBs), such as Otu1, can trim the polyubiquitin chain on a substrate and prevent its degradation.

Degradation of specific subunits from larger complexes. A: The cell cycle regulator Sic1 binds to a cyclin dependent kinase (cdk)-cyclin complex and inhibits its activity. During the cell cycle, Sic1 is degraded in a process that leaves cdk-cyclin intact despite the fact that both Sic1 and cyclin contain unstructured regions that can serve as initiation sites. Sic1 could be degraded first because its disordered region is a more effective initiation site in this complex. B: Viral infectivity factor (Vif) targets the antiretroviral cytidine deaminase APOBEC3G (A3G) for degradation. Polyubiquitinated Vif can mediate the degradation of lysine-less A3G that cannot be ubiquitinated. This could occur by trans targeting, where the ubiquitinated Vif acts as an adaptor protein to bring A3G to the proteasome. Degradation begins when A3G is engaged by the proteasome at its initiation site.
Similar articles
-
ATP-Dependent inactivation and sequestration of ornithine decarboxylase by the 26S proteasome are prerequisites for degradation.Mol Cell Biol. 1999 Oct;19(10):7216-27. doi: 10.1128/MCB.19.10.7216. Mol Cell Biol. 1999. PMID: 10490656 Free PMC article.
-
Identification of Caspase-6-mediated processing of the valosin containing protein (p97) in Alzheimer's disease: a novel link to dysfunction in ubiquitin proteasome system-mediated protein degradation.J Neurosci. 2010 Apr 28;30(17):6132-42. doi: 10.1523/JNEUROSCI.5874-09.2010. J Neurosci. 2010. PMID: 20427671 Free PMC article.
-
Identification of the Cdc48•20S proteasome as an ancient AAA+ proteolytic machine.Science. 2012 Aug 17;337(6096):843-6. doi: 10.1126/science.1224352. Epub 2012 Jul 26. Science. 2012. PMID: 22837385 Free PMC article.
-
Ubiquitin recognition by the proteasome.J Biochem. 2017 Feb 1;161(2):113-124. doi: 10.1093/jb/mvw091. J Biochem. 2017. PMID: 28069863 Review.
-
Recognition of Client Proteins by the Proteasome.Annu Rev Biophys. 2017 May 22;46:149-173. doi: 10.1146/annurev-biophys-070816-033719. Epub 2017 Mar 9. Annu Rev Biophys. 2017. PMID: 28301771 Review.
Cited by
-
Epidermal growth factor promotes protein degradation of epithelial protein lost in neoplasm (EPLIN), a putative metastasis suppressor, during epithelial-mesenchymal transition.J Biol Chem. 2013 Jan 18;288(3):1469-79. doi: 10.1074/jbc.M112.438341. Epub 2012 Nov 27. J Biol Chem. 2013. PMID: 23188829 Free PMC article.
-
Myc proteins in brain tumor development and maintenance.Ups J Med Sci. 2012 May;117(2):122-31. doi: 10.3109/03009734.2012.658975. Epub 2012 Feb 21. Ups J Med Sci. 2012. PMID: 22348395 Free PMC article.
-
Inducible Rpt3, a Proteasome Component, Knockout in Adult Skeletal Muscle Results in Muscle Atrophy.Front Cell Dev Biol. 2020 Sep 2;8:859. doi: 10.3389/fcell.2020.00859. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32984340 Free PMC article.
-
Subcellular Clearance and Accumulation of Huntington Disease Protein: A Mini-Review.Front Mol Neurosci. 2016 Apr 21;9:27. doi: 10.3389/fnmol.2016.00027. eCollection 2016. Front Mol Neurosci. 2016. PMID: 27147961 Free PMC article. Review.
-
Protein unfolding and degradation by the AAA+ Lon protease.Protein Sci. 2012 Feb;21(2):268-78. doi: 10.1002/pro.2013. Epub 2012 Jan 4. Protein Sci. 2012. PMID: 22162032 Free PMC article.
References
-
- Lee C, Schwartz MP, Prakash S, Iwakura M, Matouschek A. ATP-Dependent Proteases Degrade Their Substrates by Processively Unraveling Them from the Degradation Signal. Mol Cell. 2001;7:627–637. - PubMed
-
- Groll M, et al. Structure of 20S proteasome from yeast at 2.4 Å resolution. Nature. 1997;386:463–471. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
