

Modulation of dendritic cell maturation and function with mono- and bifunctional small interfering RNAs targeting indoleamine 2,3-dioxygenase
- PMID: 19740345
- PMCID: PMC2753901
- DOI: 10.1111/j.1365-2567.2009.03093.x
Modulation of dendritic cell maturation and function with mono- and bifunctional small interfering RNAs targeting indoleamine 2,3-dioxygenase
Abstract
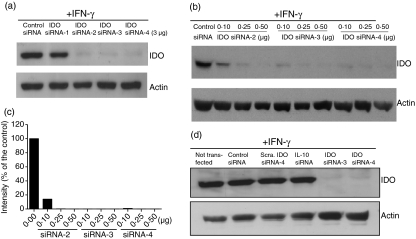
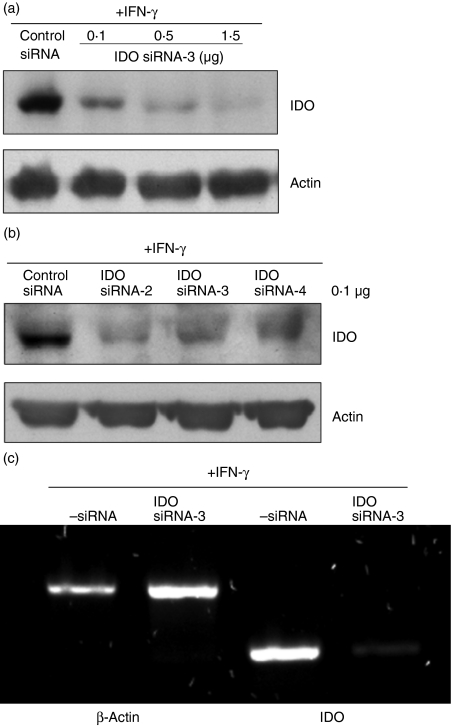
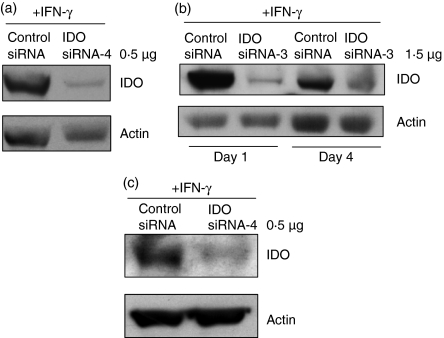
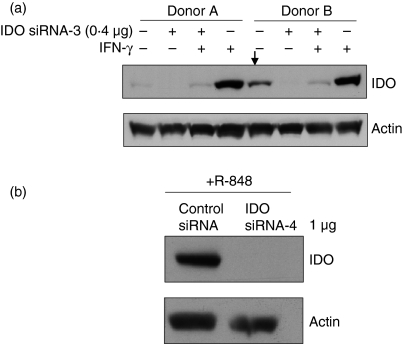
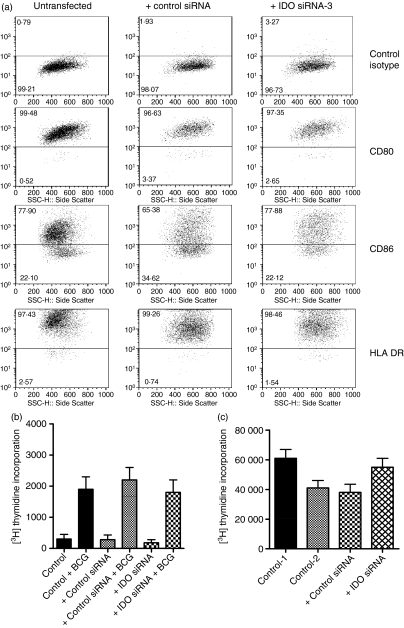
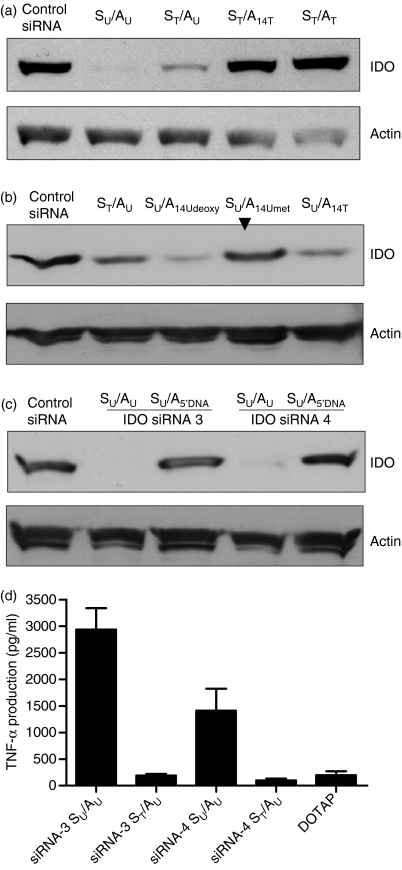
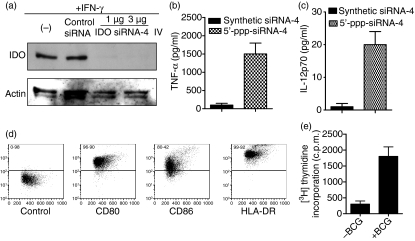
Antigen-presenting cells expressing indoleamine 2,3-dioxygenase (IDO) play a critical role in maintaining peripheral tolerance. Strategies to inhibit IDO gene expression and enhance antigen-presenting cell function might improve anti-tumour immunity. Here we have designed highly effective anti-IDO small interfering (si) RNAs that function at low concentrations. When delivered to human primary immune cells such as monocytes and dendritic cells (DCs), they totally inhibited IDO gene expression without impairing DC maturation and function. Depending on the design and chemical modifications, we show that it is possible to design either monofunctional siRNAs devoid of immunostimulation or bifunctional siRNAs with gene silencing and immunostimulatory activities. The latter are able to knockdown IDO expression and induce cytokine production through either endosomal Toll-like receptor 7/8 or cytoplasmic retinoid acid-inducible gene 1 helicase. Inhibition of IDO expression with both classes of siRNAs inhibited DC immunosuppressive function on T-cell proliferation. Immature monocyte-derived DCs that had been transfected with siRNA-bearing 5'-triphosphate activated T cells, indicating that, even in the absence of external stimuli such as tumour necrosis factor-alpha, those DCs were sufficiently mature to initiate T-cell activation. Collectively, our data highlight the potential therapeutic applications of this new generation of siRNAs in immunotherapy.
Figures
Similar articles
-
Engineering better immunotherapies via RNA interference.Hum Vaccin Immunother. 2014;10(11):3165-74. doi: 10.4161/hv.29754. Hum Vaccin Immunother. 2014. PMID: 25483669 Free PMC article. Review.
-
Impaired expression of indoleamine 2, 3-dioxygenase in monocyte-derived dendritic cells in response to Toll-like receptor-7/8 ligands.Immunology. 2008 Feb;123(2):263-71. doi: 10.1111/j.1365-2567.2007.02695.x. Epub 2007 Aug 28. Immunology. 2008. PMID: 17725606 Free PMC article.
-
Prostaglandin E2 is a key factor for monocyte-derived dendritic cell maturation: enhanced T cell stimulatory capacity despite IDO.J Leukoc Biol. 2007 Nov;82(5):1106-14. doi: 10.1189/jlb.0905519. Epub 2007 Aug 14. J Leukoc Biol. 2007. PMID: 17698915
-
Human type 1 and type 2 conventional dendritic cells express indoleamine 2,3-dioxygenase 1 with functional effects on T cell priming.Eur J Immunol. 2021 Jun;51(6):1494-1504. doi: 10.1002/eji.202048580. Epub 2021 Mar 22. Eur J Immunol. 2021. PMID: 33675038 Free PMC article.
-
T cell regulatory plasmacytoid dendritic cells expressing indoleamine 2,3 dioxygenase.Handb Exp Pharmacol. 2009;(188):165-96. doi: 10.1007/978-3-540-71029-5_8. Handb Exp Pharmacol. 2009. PMID: 19031026 Free PMC article. Review.
Cited by
-
Cancer Immunotherapy: Silencing Intracellular Negative Immune Regulators of Dendritic Cells.Cancers (Basel). 2019 Jan 17;11(1):108. doi: 10.3390/cancers11010108. Cancers (Basel). 2019. PMID: 30658461 Free PMC article. Review.
-
Releasing the Immune System Brakes Using siRNAs Enhances Cancer Immunotherapy.Cancers (Basel). 2019 Feb 3;11(2):176. doi: 10.3390/cancers11020176. Cancers (Basel). 2019. PMID: 30717461 Free PMC article. Review.
-
Engineering better immunotherapies via RNA interference.Hum Vaccin Immunother. 2014;10(11):3165-74. doi: 10.4161/hv.29754. Hum Vaccin Immunother. 2014. PMID: 25483669 Free PMC article. Review.
-
RNA Interference-Induced Innate Immunity, Off-Target Effect, or Immune Adjuvant?Front Immunol. 2017 Mar 23;8:331. doi: 10.3389/fimmu.2017.00331. eCollection 2017. Front Immunol. 2017. PMID: 28386261 Free PMC article. Review.
-
The interaction of innate immune and adaptive immune system.MedComm (2020). 2024 Sep 15;5(10):e714. doi: 10.1002/mco2.714. eCollection 2024 Oct. MedComm (2020). 2024. PMID: 39286776 Free PMC article. Review.
References
-
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–52. - PubMed
-
- Munn DH, Zhou M, Attwood JT, Bondarev I, Conway SJ, Marshall B, Brown C, Mellor AL. Prevention of allogeneic fetal rejection by tryptophan catabolism. Science. 1998;281:1191–3. - PubMed
-
- Munn DH, Mellor AL. IDO expression by dendritic cells: tolerance and tryptophan catabolism. Nat Rev Immunol. 2004;4:762–74. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
