

MicroRNA-208a is a regulator of cardiac hypertrophy and conduction in mice
- PMID: 19726871
- PMCID: PMC2735902
- DOI: 10.1172/JCI36154
MicroRNA-208a is a regulator of cardiac hypertrophy and conduction in mice
Abstract
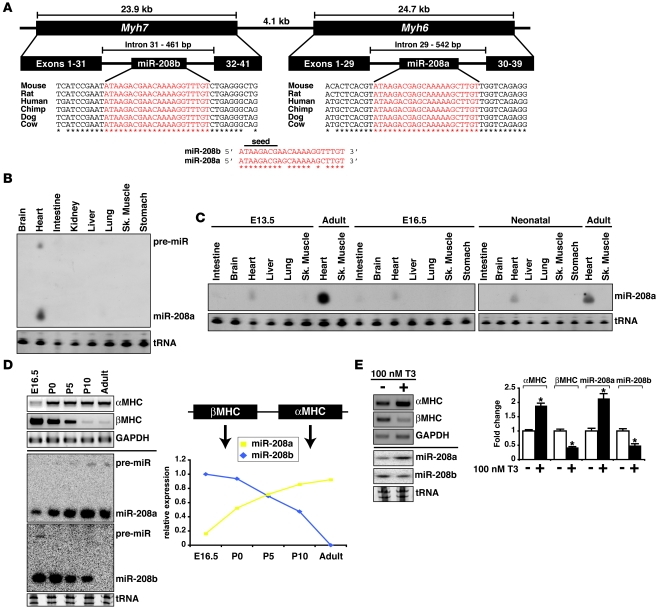
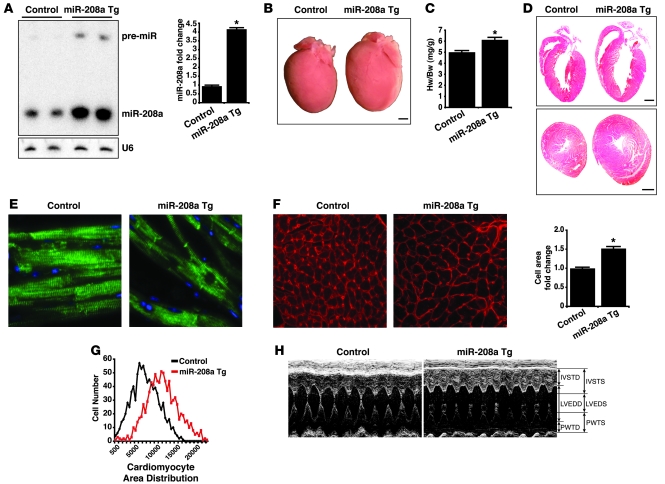
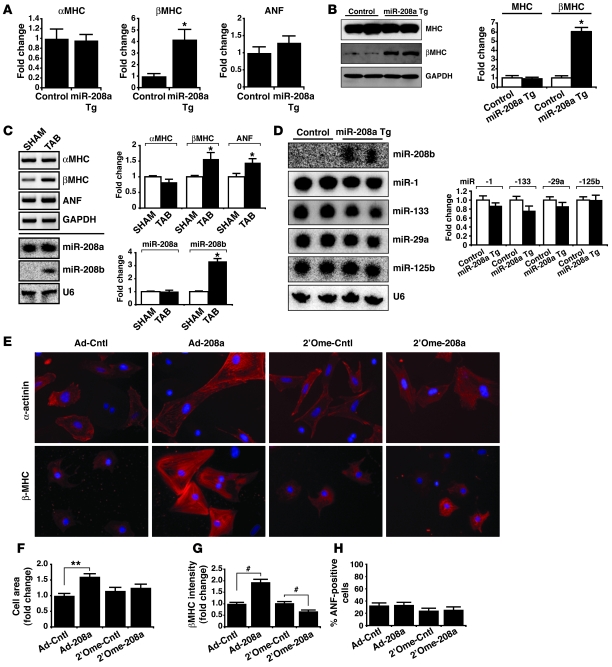
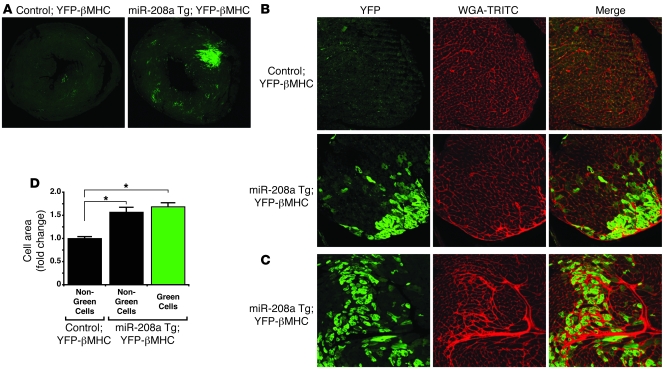
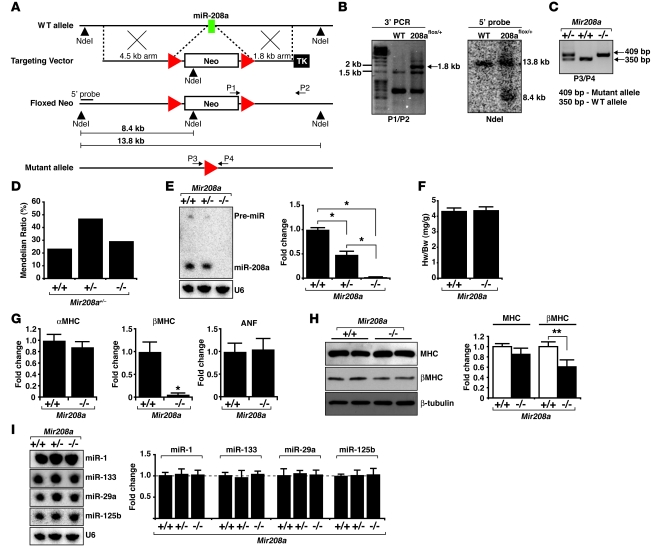
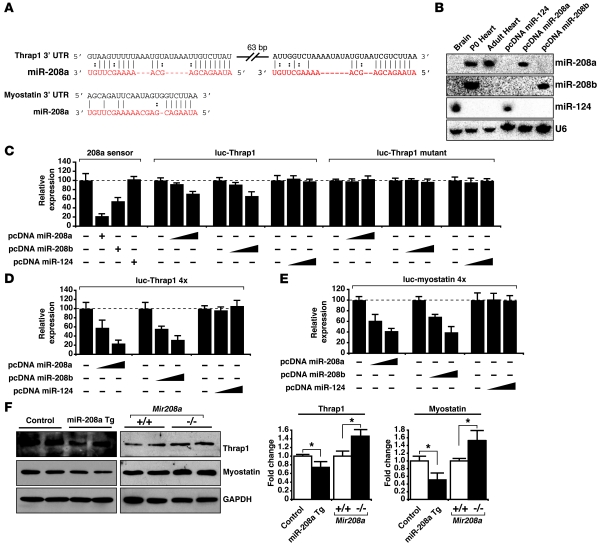
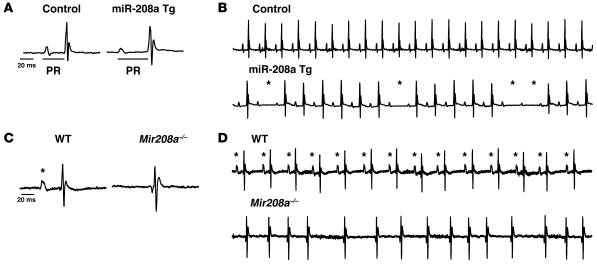
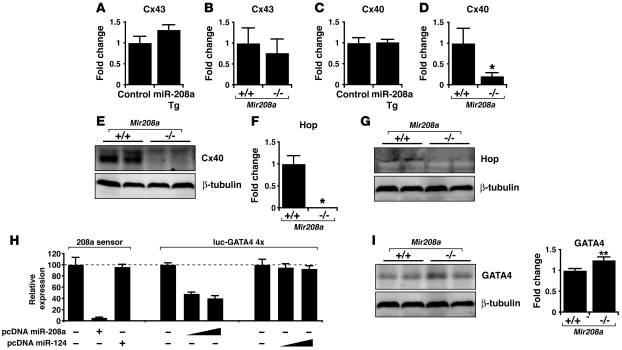
MicroRNAs (miRNAs) are a class of small noncoding RNAs that have gained status as important regulators of gene expression. Here, we investigated the function and molecular mechanisms of the miR-208 family of miRNAs in adult mouse heart physiology. We found that miR-208a, which is encoded within an intron of alpha-cardiac muscle myosin heavy chain gene (Myh6), was actually a member of a miRNA family that also included miR-208b, which was determined to be encoded within an intron of beta-cardiac muscle myosin heavy chain gene (Myh7). These miRNAs were differentially expressed in the mouse heart, paralleling the expression of their host genes. Transgenic overexpression of miR-208a in the heart was sufficient to induce hypertrophic growth in mice, which resulted in pronounced repression of the miR-208 regulatory targets thyroid hormone-associated protein 1 and myostatin, 2 negative regulators of muscle growth and hypertrophy. Studies of the miR-208a Tg mice indicated that miR-208a expression was sufficient to induce arrhythmias. Furthermore, analysis of mice lacking miR-208a indicated that miR-208a was required for proper cardiac conduction and expression of the cardiac transcription factors homeodomain-only protein and GATA4 and the gap junction protein connexin 40. Together, our studies uncover what we believe are novel miRNA-dependent mechanisms that modulate cardiac hypertrophy and electrical conduction.
Figures
Similar articles
-
The emerging role of miR-208a in the heart.DNA Cell Biol. 2013 Jan;32(1):8-12. doi: 10.1089/dna.2012.1787. Epub 2012 Nov 2. DNA Cell Biol. 2013. PMID: 23121236 Review.
-
MiRNA-208a and miRNA-208b are triggered in thyroid hormone-induced cardiac hypertrophy - role of type 1 Angiotensin II receptor (AT1R) on miRNA-208a/α-MHC modulation.Mol Cell Endocrinol. 2013 Jul 15;374(1-2):117-24. doi: 10.1016/j.mce.2013.04.010. Epub 2013 Apr 26. Mol Cell Endocrinol. 2013. PMID: 23623871
-
Early dysregulation of cardiac-specific microRNA-208a is linked to maladaptive cardiac remodelling in diabetic myocardium.Cardiovasc Diabetol. 2019 Jan 29;18(1):13. doi: 10.1186/s12933-019-0814-4. Cardiovasc Diabetol. 2019. PMID: 30696455 Free PMC article.
-
A family of microRNAs encoded by myosin genes governs myosin expression and muscle performance.Dev Cell. 2009 Nov;17(5):662-73. doi: 10.1016/j.devcel.2009.10.013. Dev Cell. 2009. PMID: 19922871 Free PMC article.
-
miR-208a in Cardiac Hypertrophy and Remodeling.Front Cardiovasc Med. 2021 Dec 9;8:773314. doi: 10.3389/fcvm.2021.773314. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34957257 Free PMC article. Review.
Cited by
-
Micro-RNA-195 and -451 regulate the LKB1/AMPK signaling axis by targeting MO25.PLoS One. 2012;7(7):e41574. doi: 10.1371/journal.pone.0041574. Epub 2012 Jul 23. PLoS One. 2012. PMID: 22844503 Free PMC article.
-
Role of microRNA in metabolic shift during heart failure.Am J Physiol Heart Circ Physiol. 2017 Jan 1;312(1):H33-H45. doi: 10.1152/ajpheart.00341.2016. Epub 2016 Oct 14. Am J Physiol Heart Circ Physiol. 2017. PMID: 27742689 Free PMC article. Review.
-
A human 3' miR-499 mutation alters cardiac mRNA targeting and function.Circ Res. 2012 Mar 30;110(7):958-67. doi: 10.1161/CIRCRESAHA.111.260752. Epub 2012 Feb 28. Circ Res. 2012. PMID: 22374132 Free PMC article.
-
Extracellular vesicle-derived miRNAs improve stem cell-based therapeutic approaches in muscle wasting conditions.Front Immunol. 2022 Nov 14;13:977617. doi: 10.3389/fimmu.2022.977617. eCollection 2022. Front Immunol. 2022. PMID: 36451814 Free PMC article.
-
Glycoproteomics Reveals Decorin Peptides With Anti-Myostatin Activity in Human Atrial Fibrillation.Circulation. 2016 Sep 13;134(11):817-32. doi: 10.1161/CIRCULATIONAHA.115.016423. Epub 2016 Aug 24. Circulation. 2016. PMID: 27559042 Free PMC article.
References
-
- Filipowicz W., Bhattacharyya S.N., Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat. Rev. Genet. 2008;9:102–114. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
