

Modification of high density lipoprotein by myeloperoxidase generates a pro-inflammatory particle
- PMID: 19726691
- PMCID: PMC2781481
- DOI: 10.1074/jbc.M109.047605
Modification of high density lipoprotein by myeloperoxidase generates a pro-inflammatory particle
Abstract
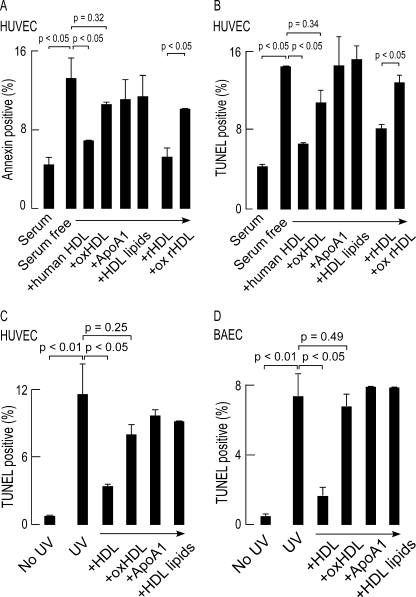
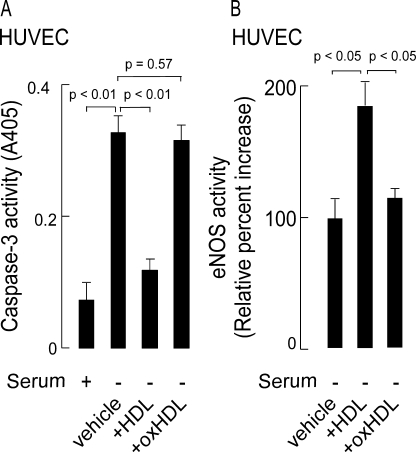
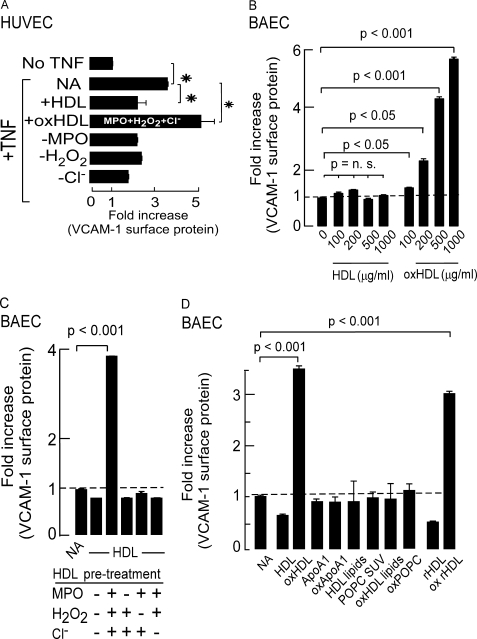
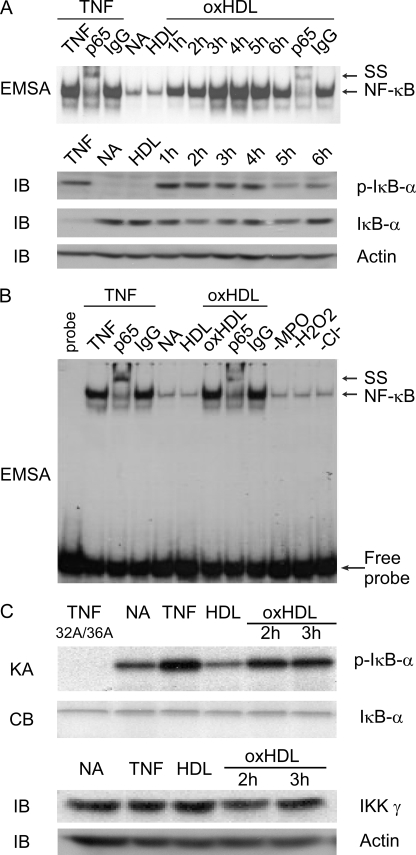
High density lipoprotein (HDL) is the major atheroprotective particle in plasma. Recent studies demonstrate that myeloperoxidase (MPO) binds to HDL in vivo, selectively targeting apolipoprotein A1 (apoA1) of HDL for oxidative modification and concurrent loss in cholesterol efflux and lecithin cholesterol acyl transferase activating activities, generating a "dysfunctional HDL" particle. We now show that (patho)physiologically relevant levels of MPO-catalyzed oxidation result in loss of non-cholesterol efflux activities of HDL including anti-apoptotic and anti-inflammatory functions. One mechanism responsible is shown to involve the loss of modified HDL binding to the HDL receptor, scavenger receptor B1, and concurrent acquisition of saturable and specific binding to a novel unknown receptor independent of scavenger receptors CD36 and SR-A1. HDL modification by MPO is further shown to confer pro-inflammatory gain of function activities as monitored by NF-kappaB activation and surface vascular cell adhesion molecule levels on aortic endothelial cells exposed to MPO-oxidized HDL. The loss of non-cholesterol efflux activities and the gain of pro-inflammatory functions requires modification of the entire particle and can be recapitulated by oxidation of reconstituted HDL particles comprised of apoA1 and nonoxidizable phosphatidylcholine species. Multiple site-directed mutagenesis studies of apoA1 suggest that the pro-inflammatory activity of MPO-modified HDL does not involve methionine, tyrosine, or tryptophan, oxidant-sensitive residues previously mapped as sites of apoA1 oxidation within human atheroma. Thus, MPO-catalyzed oxidation of HDL results not only in the loss of classic atheroprotective reverse cholesterol transport activities of the lipoprotein but also both the loss of non-cholesterol efflux related activities and the gain of pro-inflammatory functions.
Figures

Similar articles
-
An abundant dysfunctional apolipoprotein A1 in human atheroma.Nat Med. 2014 Feb;20(2):193-203. doi: 10.1038/nm.3459. Epub 2014 Jan 26. Nat Med. 2014. PMID: 24464187 Free PMC article.
-
Myeloperoxidase, paraoxonase-1, and HDL form a functional ternary complex.J Clin Invest. 2013 Sep;123(9):3815-28. doi: 10.1172/JCI67478. Epub 2013 Aug 1. J Clin Invest. 2013. PMID: 23908111 Free PMC article.
-
HDL from apoA1 transgenic mice expressing the 4WF isoform is resistant to oxidative loss of function.J Lipid Res. 2015 Mar;56(3):653-664. doi: 10.1194/jlr.M056754. Epub 2015 Jan 5. J Lipid Res. 2015. PMID: 25561462 Free PMC article.
-
Formation of dysfunctional high-density lipoprotein by myeloperoxidase.Trends Cardiovasc Med. 2005 Aug;15(6):212-9. doi: 10.1016/j.tcm.2005.06.004. Trends Cardiovasc Med. 2005. PMID: 16182131 Review.
-
Myeloperoxidase-mediated oxidation of high-density lipoproteins: fingerprints of newly recognized potential proatherogenic lipoproteins.Arch Biochem Biophys. 2006 Jan 15;445(2):245-55. doi: 10.1016/j.abb.2005.08.008. Epub 2005 Aug 31. Arch Biochem Biophys. 2006. PMID: 16171772 Review.
Cited by
-
Crosstalk between reverse cholesterol transport and innate immunity.Trends Endocrinol Metab. 2012 Apr;23(4):169-78. doi: 10.1016/j.tem.2012.02.001. Epub 2012 Mar 10. Trends Endocrinol Metab. 2012. PMID: 22406271 Free PMC article. Review.
-
Oxidation of methionine residues in human apolipoprotein A-I generates a potent pro-inflammatory molecule.J Biol Chem. 2019 Mar 8;294(10):3634-3646. doi: 10.1074/jbc.RA118.005663. Epub 2019 Jan 11. J Biol Chem. 2019. PMID: 30635405 Free PMC article.
-
Good Cholesterol Gone Bad? HDL and COVID-19.Int J Mol Sci. 2021 Sep 22;22(19):10182. doi: 10.3390/ijms221910182. Int J Mol Sci. 2021. PMID: 34638523 Free PMC article. Review.
-
Mechanisms underlying adverse effects of HDL on eNOS-activating pathways in patients with coronary artery disease.J Clin Invest. 2011 Jul;121(7):2693-708. doi: 10.1172/JCI42946. Epub 2011 Jun 23. J Clin Invest. 2011. PMID: 21701070 Free PMC article.
-
Effects of native and myeloperoxidase-modified apolipoprotein a-I on reverse cholesterol transport and atherosclerosis in mice.Arterioscler Thromb Vasc Biol. 2014 Apr;34(4):779-89. doi: 10.1161/ATVBAHA.113.303044. Epub 2014 Jan 9. Arterioscler Thromb Vasc Biol. 2014. PMID: 24407029 Free PMC article.
References
-
- Fielding C. J., Fielding P. E. (1995) J. Lipid Res. 36, 211–228 - PubMed
-
- Tall A. R. (1998) Eur. Heart. J. 19, (Suppl. A) A31–A35 - PubMed
-
- Barter P. J., Nicholls S., Rye K. A., Anantharamaiah G. M., Navab M., Fogelman A. M. (2004) Circ. Res. 95, 764–772 - PubMed
-
- Hessler J. R., Robertson A. L., Jr., Chisolm G. M., 3rd (1979) Atherosclerosis 32, 213–229 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
